Growing season water balance of an inner alpine Scots pine (Pinus sylvestris L.) forest
iForest - Biogeosciences and Forestry, Volume 11, Issue 4, Pages 469-475 (2018)
doi: https://doi.org/10.3832/ifor2626-011
Published: Jul 02, 2018 - Copyright © 2018 SISEF
Research Articles
Abstract
We estimated components of the water cycle of a 150-year-old Pinus sylvestris forest in an inner Alpine dry valley of the Tyrol, Austria throughout five growing seasons. Forest canopy transpiration (TC) was measured by sap flow measurements scaled to the stand canopy level. Estimates of understory transpiration and forest floor evaporation (ETU) were derived from the soil water budget method, while interception (I) was modelled. Growing season cumulative evapotranspiration (ET = TC + ETU + I) varied between 256 and 322 mm or 51 to 79% of the growing season precipitation. The contribution of TC, ETU, and I to ET were 33, 40 and 27% respectively. Although these values of each layer (evapo)-transpiration are in good agreement with studies carried out in other European Scots pine forests, our estimated growing season total forest water use (Ttot = Tc + ETu) of 200-244 mm is at the lower end of values reported for coniferous forest ecosystems, and thus reflects an adaptation to the low shallow soil water availability. We conclude that Scots pine forests in inner alpine dry valleys are able to cope with high evaporative demand, even when shallow soil water availability is limited.
Keywords
Forest Water Balance, Scots Pine, Dry Inner Alpine Valley, Evapotranspiration, Interception, Runoff
Introduction
Scots pine (Pinus sylvestris L.) is a widespread conifer species across Eurasia. In Europe, P. sylvestris can be found from the boreal north (70 °N) in Scandinavia to the Mediterranean basin (37 °N) in southern Spain ([27]) and it forms open stands in inner alpine dry valleys. As a consequence of climate change, changes of the ecosystem water balance are expected to occur especially in open Pinus sylvestris forests in inner- alpine dry valleys in Italy, Switzerland and Austria, which are characterized by high summer temperatures, low precipitation and limited soil water availability ([40], [25], [37]), and thus are considered to be sensitive to climate change ([28], [30]).
The water balance of a forest ecosystem is determined by the water input through precipitation (P). Some of the P adheres on the above-ground vegetation and evaporates before reaching the ground - the so-called interception (I). The remaining P reaching the soil surface runs off (RS, surface runoff), infiltrates into the soil, and percolates through the water table (RD, deep seepage), finally leaving the system as base flow. The amount of water which is held against gravitational forces within the soil matrix can be taken up by the plant roots and is finally transpired by the canopy (TC) and evapotranspirates from the forest floor (ETU, soil evaporation and understory transpiration) into the atmosphere. Thus, over short periods there are also changes in the soil water content (ΔW). All the components involved are given in mm (= kg m-2), and the water balance equation of a forest ecosystem reads (eqn. 1):
in which all terms, except ΔW, are flux densities.
As it is frequently very difficult to measure all the components of eqn. 1, the water balance equation (eqn. 1) can be simplified as (eqn. 2):
where ET is the sum of all water fluxes from the forest to the atmosphere (TC + ETU + I) and R (total runoff) is the sum of deep seepage and surface runoff.
Although data on the water balance of Scots pine forests have been reported by several authors ([34], [12], [15]), we are not aware of any study focusing on the water balance of Scots pine stands in inner Alpine dry valleys. Therefore, it was the aim of this paper to quantify and describe the main terms of the hydrological cycle in an inner alpine dry Scots pine forest and the parameters that affect total forest water use (Ttot = TC + ETU). One particular task was to quantify the contribution of TC, ETU, and I on ET, by applying scaled sap flow rates, estimates of ETU using the soil water budget method and estimates of I derived from precipitation measurements and literature data. To achieve this goal, we monitored environmental data (air temperature, T; relative humidity, RH; global radiation, GR; precipitation, P; soil moisture, θ; and sap flow density, Qs) during the growing seasons (April 1 - October 31) of 2007, 2008, 2009, 2011, and 2012. We, however, were not able to take measurements during the 2010 growing season due to a lack of funding.
Material and methods
Study site
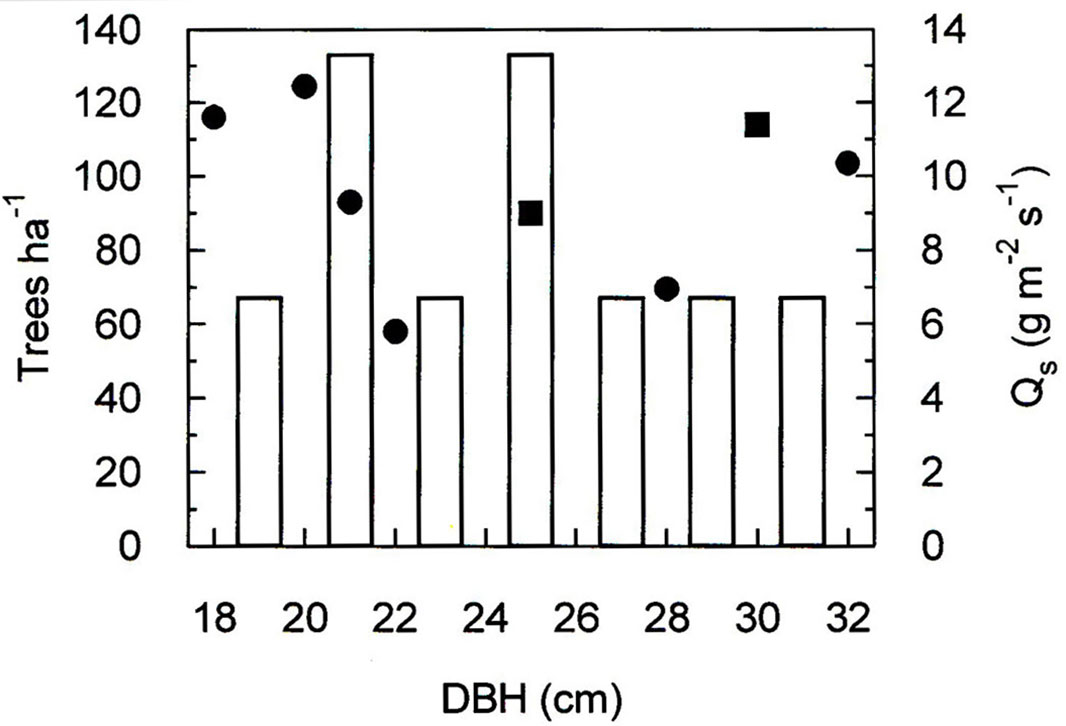
The study was carried out in an open Spring heath-Scots pine forest (Erico-Pinetum typicum - [9]) growing on a postglacial rock-slide area, situated in the montane belt (750 m a.s.l.) within the inner Alpine dry Inn valley (Tyrol, Austria - 47° 14′ 00″ N, 10° 50′ 20″ E). The study site is south exposed (40° inclination - [37]) and pioneer vegetation (mainly Erica carnea and Sesleria varia) prevails in the ground flora. At the time of the study (2007-2012), the trees were 150 ± 30 years old and their height ranged between 4 and 5 m. The stand density was 600 trees ha-1, with a canopy coverage of 30%, a leaf area index (LAI) of 0.55 ([25]), and a basal area of 21.8 m2 ha-1. The tree diameter at breast height (DBH) averaged 24.5 ± 5.1 cm and varied between 18 and 32 cm (Fig. 1).
Fig. 1 - Stocking density (trees ha-1) at the study site (open bars) and daily maximum sap flow density (Qs) of the selected study trees during the growing seasons 2007 - 2009 (squares) and 2011 - 2012 (circles) in relation to diameter at breast height (DBH).
The site is characterized by a continental climate with a mean annual temperature of 7.3 °C and a mean annual precipitation of 718 mm (long-term mean 1911-2012 at Ötz, 812 m a.s.l., 5 km south of the study site), and spring drought, occurring every third year at average ([11]). Furthermore, the water balance is strained by a low water holding capacity of the protorendzina, i.e., a rendzic leptosol (O-A-C profile) according to the World Base for Soil Resources classification system ([10]). The A horizon reaches a depth of 10 cm and consists of coarse-textured materials. The texture is dominated by the sand (54%) and the silt fractions (44%) with almost no clay (2%). According to the water retention curve obtained for the study site ([37]), the water holding capacity is 42 mm at field saturation. The corresponding values for field capacity (-0.033 MPa, sensu [2]) is 23 mm, and the wilting point (-1.5 MPa) is reached at 7 mm.
Environmental measurements
Air temperature (T), relative humidity (RH), global radiation (GR), and precipitation (P) were monitored 2 m above the ground in an open, non-vegetated ridge in order to minimize interception loss. During the study periods 2007-2009 all these environmental data were recorded with an automatic weather station (ONSET®, Pocasset, MA, USA). In 2011 and 2012 T, RH (HMP45C®, Campbell Scientific, Shepshed, UK), GR (SP-Lite®), and P (ARG100®) were recorded with a Campbell CR10X® data logger (Campbell, Scientific, Shepshed, UK). Both data loggers were programmed to record 30-min averages of all the environmental data sampled every minute during the growing seasons (April 1 - October 31) of 2007, 2008, 2009, 2011, and 2012. Throughout the same periods, soil moisture (θ, m3 m-3) was continuously monitored in 5-10 cm soil depth at three sites with capacitive soil moisture sensors (Cyclobis, proprietary development, University Innsbruck, Austria). The measuring interval of these sensors was 30 min, and due to small-scale variability of the soil structure ([25]) values of the three soil moisture sensors were averaged. Finally, T, RH, GR, and θ were expressed as daily means, while P was summed up to daily totals (48 values per day each).
Radiation and temperature data obtained 2 m above the ground were used to calculate daily potential evapotranspiration (PET, mm day-1) using the following formula ([33], see also [18] - eqn. 3):
where Rg is the daily sum of global radiation (MJ m-2 day-1) and Ta is the daily mean air temperature.
Shallow soil water deficit was quantified as relative extractable shallow soil water (REW, dimensionless) and was calculated as follows ([14], [34]) - eqn. 4):
where Wd is the actual soil water content (mm) on day d, Wmin is the minimum soil water content at the permanent wilting point, and Wfc is soil water content at field capacity. For P. sylvestris trees in nearby more mesic Erico-Pinetumtypicum forest, an REW ≤ 0.4 has been shown to limit crown conductance ([20]).
Determination of water fluxes
Sap flow density and estimation of canopy transpiration
Sap flow density (Qs) was monitored with thermal dissipation sensors according to Granier ([13]) by battery-operated sap flow systems (M1Sapflow System®, PROSA-LOG UP, Umweltanalytische Produkte GmbH, Cottbus, Germany). In each study tree (two in 2007-2009 and six in 2011-2012) one 20 mm long sensor was installed into the outer xylem (0-20 mm from the cambium) at breast height. Accounting for variations of Qs across the cross section of the trunk ([5]), sensors were also installed in the next 20 mm thick xylem band (20-40 mm sapwood depth, termed “inner xylem”) in two of the selected study trees in 2011 and 2012. The two probes of each sensor were installed vertically 15 cm apart on the north facing side of the 1.3 m above the ground. The upper probe was heated continuously, while the lower one was unheated, remaining at trunk temperature for reference. The temperature difference between the upper heated and the lower reference probe was recorded every 30 min. The sensors were shielded with a thick aluminium-faced foam cover to prevent exposure to rain, and to avoid physical damage and thermal influences from radiation.
For each sensor, Qs (g m-2 s-1) was calculated using the original calibration coefficients of Granier ([13]). The sensors in the outer and the inner xylem covered most of the active sapwood which in our study trees had an average thickness of 41.5 ± 5.4 mm. In the study trees, Qs did not vary considerably across sapwood depth (P > 0.5). In 2011 Qs averaged 3.9 ± 2.6 and 4.0 ± 2.8 g m-2 s-1 in the outer (0-20 mm) and the inner xylem (20-40 mm), respectively. The corresponding values for 2012 were 4.8 ± 1.7 and 4.8 ± 3.01 g m-2 s-1, respectively. Thus, we assumed uniform Qs across sapwood depth and our installation sampled 100% of the whole water flow in this conifer. Since Qs was not correlated with DBH at our study site (R2 = 0.001, P = 0.45 - Fig. 1), canopy transpiration (Tc) was determined as follows ([39] - eqn. 5):
where Qs-mean is the average sapwood density if the sample trees and As-stand is the cumulative sapwood area per ground surface area (m2 m-2). A stand specific sapwood area of 10.5 cm2 m-2 ([37]) was used for up-scaling. Scaling up on this method may be applied in uniform stands with a limited range in tree size and a high individual variation of tree specific Qs ([5]) due to environmental factors (e.g., drought) as it was the case at our study site. Moreover, there is also evidence that when using 2 or 6 sample trees, the scaling error for estimates of Qs-mean is usually less than 12 and 7%, respectively ([7]).
Estimation of forest floor evapotranspiration
Forest floor evapotranspiration (ETU) could not be neglected at our study plot because of the low canopy coverage (30%). Daily ETU was estimated from shallow soil water content measurements, according to the soil water budget approach ([18]). In our experimental plot an important amount of P leaches to deeper soil layers via the macropores (gravitational water) after refilling the shallow soil (personal unpublished observations). Therefore, we assumed that ETU solely absorbed shallow soil water ([35]) and that ETU is mainly dependent on evaporative demand reaching the forest floor ([14]). Because of P inputs, water movement within the soil profile, and the ETU influence on θ, we eliminated all days with rain and also up to two days following heavy rain events (> 25 mm) for estimating ETU from ΔW measurements. Thus, ETU was calculated as follows (eqn. 6):
where ΔW (mm) is the change in shallow soil water content between two consecutive days. This allowed to use approximately 65% of growing season days for estimating ETU from the soil water budget approach. Missing ETU values were calculated using a regression between ETU and PET (R²= 0.92; P < 0.001), as there is evidence that the forest floor vegetation is poorly coupled to the atmosphere ([16], [1]).
Interception and runoff
Canopy interception (I) was determined according to measurements of Brechtel ([3]) carried out in a 120-year-old P. sylvestris forest, with a canopy coverage of 30%. Basically, we assumed I to be 23% of bulk precipitation for rainfall events < 7 mm and 10% of bulk precipitation for rainfall events > 7 mm. Finally, runoff (R) was calculated as the residual of eqn. 2.
Data analysis
In this study, we also emphasized the comparison of total forest water use (Ttot) as affected by shallow soil water availability (non-limiting vs. limiting shallow soil water availability). Single variable analyses were used to examine the response of Ttot to GR, VPD and W. While correlations of Ttot with GR and W were obtained by linear regression analysis, the relationship of Ttot with VPD was analysed using the following exponential saturation function (eqn. 7):
where a is a fitting parameter and TtotMAX is the maximum daily mean total forest water use. In addition, we used a multiple linear regression model including GR, VPD, and W, as explanatory variables for Ttot. All the analyses were performed using the software package SPSS® ver. 16.0 for Windows (SPSS Inc., Chicago, USA) and nonlinear curve fits were performed using Fig.P® for Windows (FigP Software Corporation, Hamilton, ON, Canada).
Results
Environmental conditions
Seasonal patterns of environmental conditions obtained during the growing seasons (April 1 - October 31) of 2007, 2008, 2009, 2011, and 2012 were representative for the climatic conditions at the study site, which is characterized by a dry and cool spring and a wet and warm summer. Due to differences in cloud cover, global radiation (GR) varied between 6.0 W m-2 (8 June 2011) and 339.6 W m-2 (22 May 2008 - Fig. 2), averaging 177 W m-2 in 2007, 173 W m-2 in 2008, 171 W m-2 in 2009, 168 W m-2 in 2011, and 174 W m-2 in 2012. Daily mean air temperature (T) was 14.6 °C in 2007, 14.0 °C in 2008, 14.9 °C in 2008, 14.6 °C in 2011 and 15.0 °C in 2012 and varied between 1.1 °C on October 21, 2007 and 26.3 °C on June 1, 2012 (Fig. 2). Daily mean vapour pressure deficit (VPD) was 0.53 kPA in 2007, 0.50 kPA in 2008 and in 2009, 0.62 kPA in 2011 and 0.65 kPa in 2012. Daily mean VPD approached zero on rainy days and reaching values up to 2.0 kPa on warm and sunny days during late spring and summer (data not shown).
Fig. 2 - Seasonal course of daily mean global radiation (GR), air temperature (T), daily sum of precipitation (P) and shallow soil water content (W) in 0-10 cm soil depth during the growing seasons (April 1 - October 31) 2007, 2008, 2009, 2011, and 2012. The fine solid line indicates shallow soil water deficit.
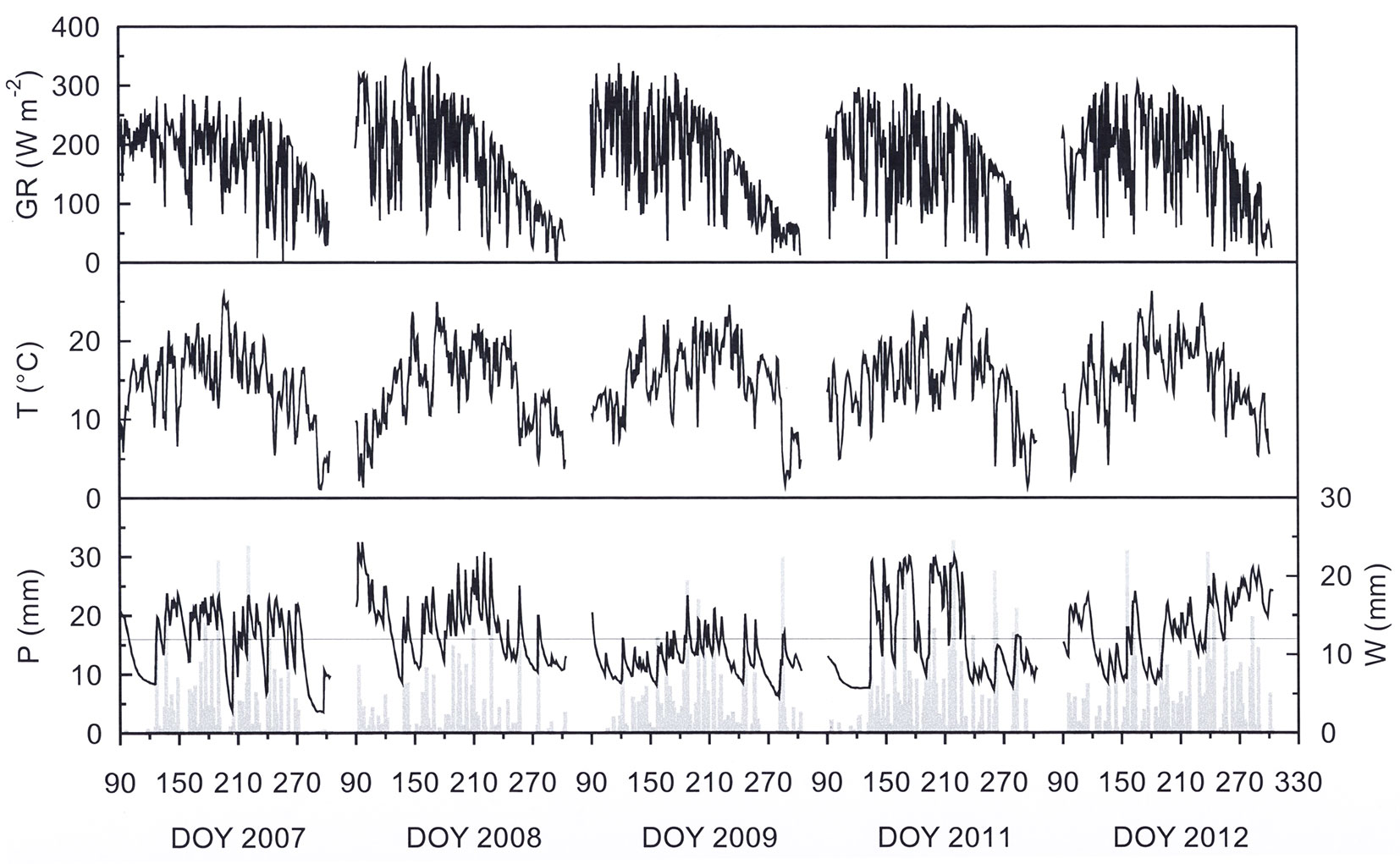
Precipitation (P) varied considerably during the course of this investigation (Fig. 2). Of the five years examined, 2008 was the driest growing season (325 mm) and 2012 was the wettest growing season with a total of 589 mm (Tab. 1). Shallow soil water content (W) varied between a maximum of 24 mm on April 3, 2008 and a minimum of 3 mm on October 23, 2007 (Fig. 2). Average W over the growing seasons 2007, 2008, 2009, 2011, and 2012 was 12, 13, 10, 12 and 13 mm, respectively. Based on an REW of 0.4 (W = 12 mm), soil water deficit characterized 24, 14, 77, 50, and 32% of the growing seasons in 2007, 2008, 2009, 2011, and 2012, respectively (Fig. 2).
Tab. 1 - Growing season (April 1 - October 31) totals of precipitation (P, mm), canopy transpiration (TC, mm), forest floor evapotranspiration (ETU, mm), total forest water use (Ttot = TC + ETU, mm), interception (I, mm), total actual evapotranspiration (ET = TC + ETU + I, mm), change in the soil water content (ΔW, mm), runoff (R, mm), the ratio of each layer (evapo)-transpiration to ET, as well as evapotranspiration (ET/P) and runoff (R/P) normalized to P. (CV): coefficient of variation.
| Year | P | T C | ET U | T tot | I | ET | ΔW | R | TC/ET | ETU/ET | Ttot/ET | I/ET | ET/P | R/P |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2007 | 398 | 90 | 118 | 208 | 65 | 273 | -8 | 133 | 0.33 | 0.43 | 0.76 | 0.24 | 0.69 | 0.33 |
| 2008 | 325 | 116 | 86 | 202 | 55 | 256 | -6 | 75 | 0.45 | 0.34 | 0.79 | 0.21 | 0.79 | 0.23 |
| 2009 | 437 | 109 | 135 | 244 | 78 | 322 | -7 | 123 | 0.34 | 0.42 | 0.76 | 0.24 | 0.74 | 0.28 |
| 2011 | 574 | 78 | 122 | 200 | 96 | 295 | -2 | 281 | 0.26 | 0.41 | 0.68 | 0.32 | 0.51 | 0.49 |
| 2012 | 589 | 85 | 126 | 211 | 100 | 310 | 7 | 273 | 0.27 | 0.41 | 0.68 | 0.32 | 0.53 | 0.46 |
| Average | 465 | 95 | 117 | 213 | 79 | 291 | -3 | 177 | 0.33 | 0.40 | 0.73 | 0.27 | 0.65 | 0.36 |
| CV (%) | 25 | 16 | 15 | 8 | 25 | 9 | -224 | 53 | 20 | 11 | 6 | 17 | 19 | 31 |
Seasonal water fluxes and influencing factors
Growing season cumulative evapotranspiration (ET) ranged from 256 mm (2008) to 322 mm (2009) (Tab. 1), or 51% (2011) to 79% (2008) of growing season precipitation with coefficients of variation of 9 and 19%, respectively (Tab. 1). Growing season cumulative evapotranspiration was composed of the following fluxes: 78 (2011) to 116 mm (2009) from TC, 86 mm (2008) to 135 mm (2008 and 2009) from ETU, and 55 mm (2008) to 100 mm (2012) from I (Tab. 1). Consequently, growing season total forest water use (Ttot = TC + ETU) varied between 200 mm in 2011 and 244 mm in 2009 (Tab. 1). Accordingly, for the stand Ttot/ET varied between 68 and 79% and the contribution of I to ET was 21 to 32% (Tab. 1).
The potential driving forces considered (global radiation, air temperature, vapour pressure deficit, soil water availability, and precipitation) explained little of the variability in growing season ET (all p > 0.67 - data not shown). The best predictor was the growing season mean air temperature, which in a linear regression explained 23% of the variability in growing season ET (p = 0.41). Restricting the analysis to Ttot (= TC + ETU), where most of the ET occurs (Tab. 1), improved regression statistics only slightly (all p > 0.40 - data not shown).
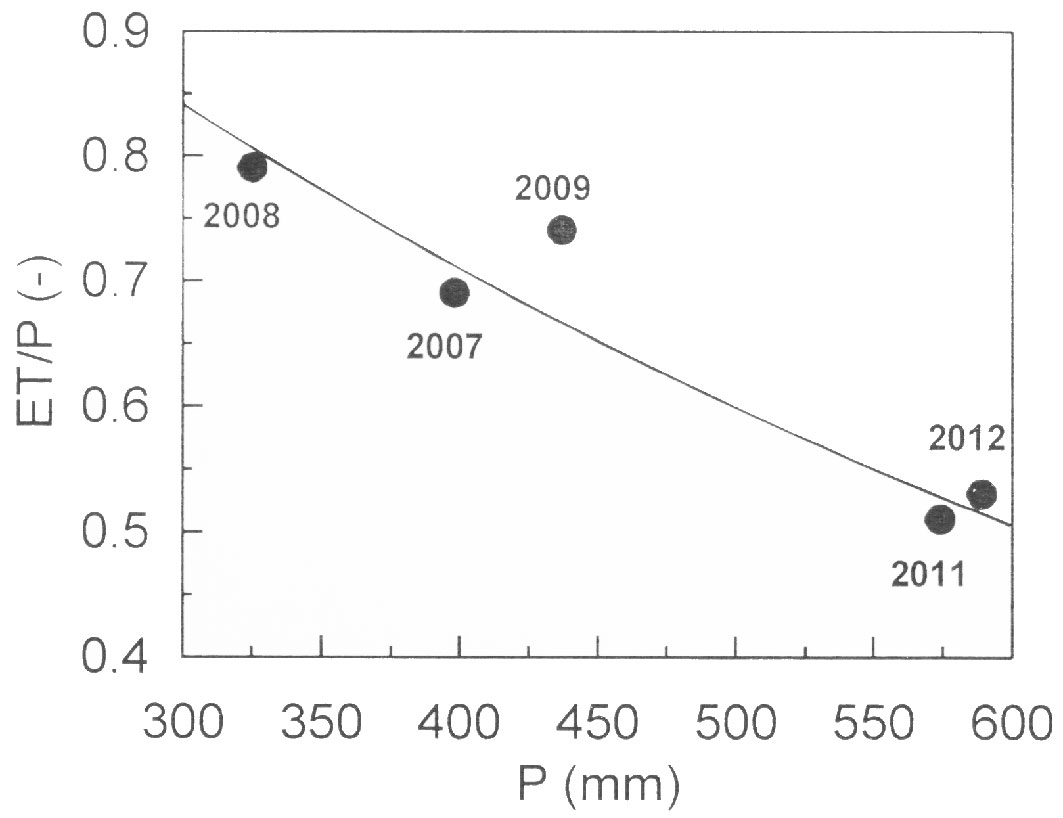
When ET was normalized with P, we found a statistically significant non-linear relationship (R² = 0.88; p < 0.001) with P (Fig. 3), indicating that the fraction of P evaporated to the atmosphere increased with decreasing P, while the runoff normalized to precipitation increased during years with ample P (Tab. 1).
Fig. 3 - Growing season evapotranspiration normalized with precipitation (ET/P) against precipitation (P). Points were fit by the exponential regression: y = 1.40· exp(-0.0017·x); R² = 0.88; p < 0.001.
Seasonal trends in daily total forest water use (Ttot) per unit ground area in general followed those of GR and T (Fig. 4, cf. Fig. 2). Ttot varied between 2.07 mm d-1 during cloudless days in summer (July 21, 2007) and 0.14 mm d-1 (October 8, 2011 and October 15, 2012) during rainy days in fall (Fig. 4). Annual growing season Ttot varied between 200 mm in 2011 and 244 mm in 2009 (Tab. 1), increased almost linearly from April throughout August and in general tended to decline gradually towards the end of the growing season throughout fall (Fig. 5).
Fig. 4 - Seasonal course of daily total forest water use (Ttot) per unit ground surface area during the growing seasons (April 1 - October 31) 2007, 2008, 2009, 2011, and 2012.
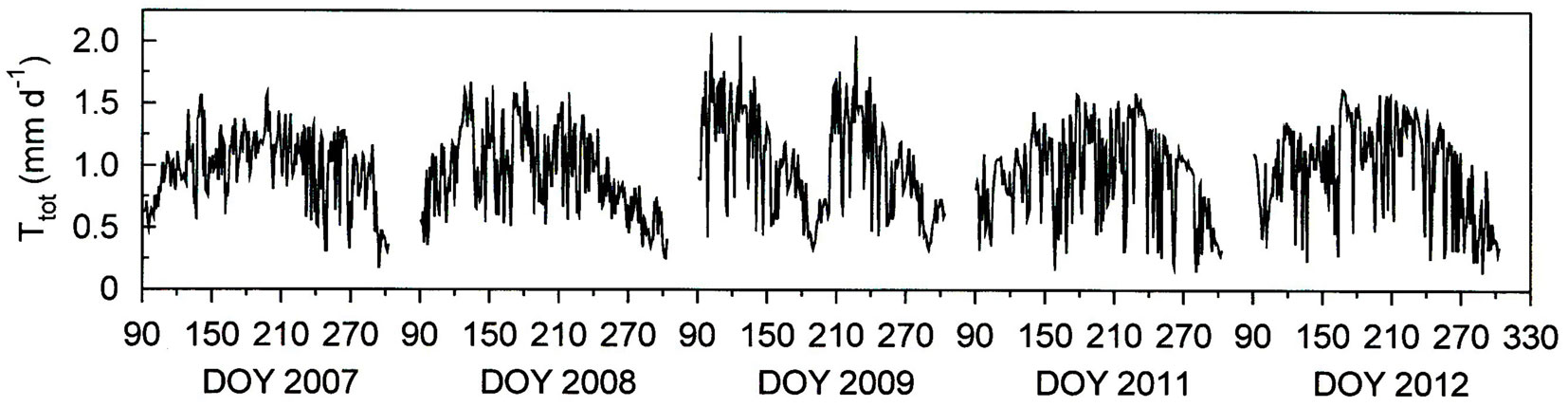
Fig. 5 - Inter-annual variation of cumulative total forest water use (Ttot) per unit ground surface area during the growing seasons (April 1 - October 31) 2007, 2008, 2009, 2011, and 2012.

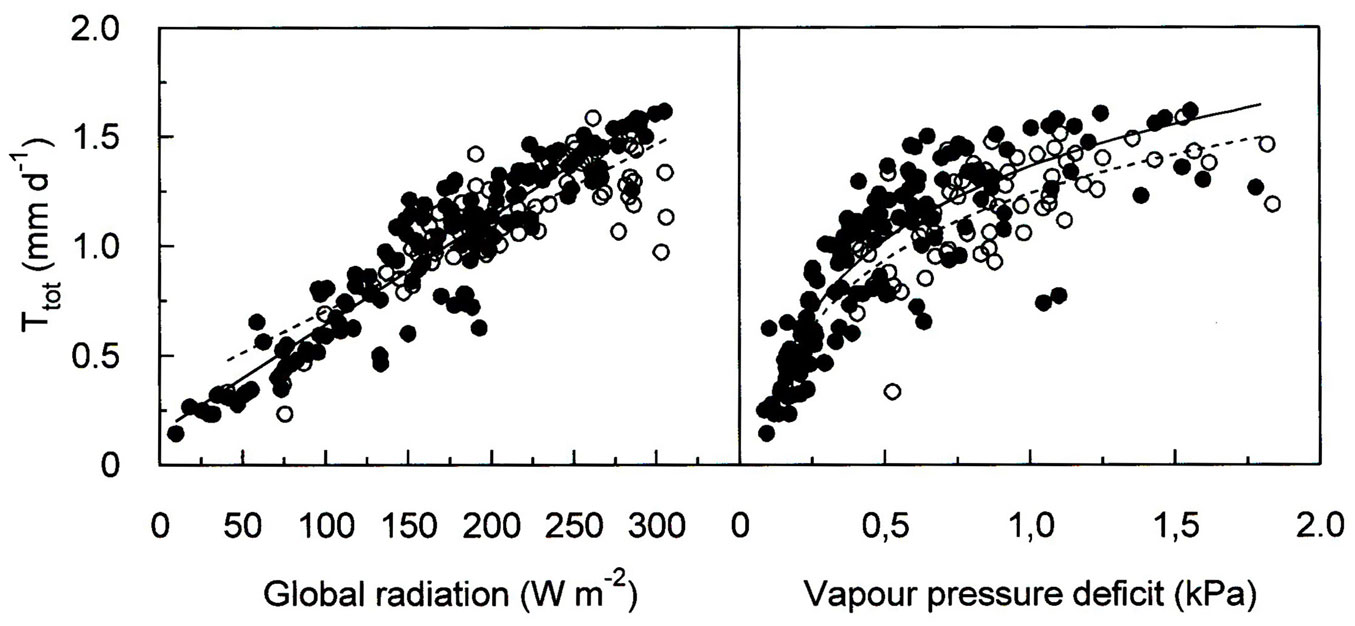
Topsoil water availability did not considerably modify the response of Ttot to GR and VPD. When examined at a daily timescale, these results generally reflected positive correlations between Ttot and both environmental factors, as shown for the growing season 2012 in Fig. 5. We obtained linear correlations between Ttot and GR at R² levels of 0.87 and 0.69 under conditions of non-limiting and limiting soil water availability (both p-values < 0.001 - Fig. 6), respectively. With respect to VPD, Ttot increased sharply at low VPD and tended to saturate at mean daily VPD values > 1.2 kPa under conditions of non-limiting shallow soil water availability and VPD values > 1.5 kPa when shallow soil water availability was limiting (Fig. 6). Vapour pressure deficit explained 74 and 63% (both p-values > 0.001) of the variation in Ttot under conditions of non-limiting and limiting shallow soil water availability, respectively. We, however, did not obtain an effect of shallow soil water availability on Ttot (p = 0.24, data not sown). A multiple linear regression model including GR, VPD and W explained 82% of the observed variability in Ttot (p < 0.001). In addition, the multiple linear regression analysis also clearly indicated that GR (β-coefficient = 0.79) had a greater effect on Ttot than VPD (β-coefficient = 0.20), while the effect of W on Ttot was nil (β-coefficient = 0.04).
Fig. 6 - Daily total forest water use (Ttot) as a function of global radiation (GR, left) and vapour pressure deficit (VPD, right) under non-limiting (solid symbols and solid line) and limiting (open symbols and dotted line) shallow soil water content. Points were fit by linear and exponential saturation functions for GR and VPD, respectively. Global radiation: non-limiting y = 0.005·GR + 0.15, R2 = 0.87; limiting W: y = 0.004·GR + 0.32, R2 = 0.69. Vapour pressure deficit: non-limiting W: y = [1-(exp(2.03·VPD))]·1.63, R2 = 0.74; limiting W: y = (1-(exp(1.59·VP)))·1.63, R2 = 0.63. All p < 0.001.
Discussion
In the present study we focused on the growing season water balance of a 150-year-old P. sylvestris forest in an inner Alpine dry valley in Tyrol, Austria, where soil water availability is limited by a low water holding capacity of the shallow soil, and where trees ([20]) and understory vegetation do not have access to groundwater. Shallow soil water reserves demonstrated clear seasonal variations (Fig. 3). Based on an RWE of 0.4 (W < 12 mm) limited shallow soil water availability prevailed for 14% (36 days in 2008) to 77% (165 days in 2009) during an entire growing season. In contrast to findings of Sturm et al. ([32]) who showed that water loss of a Scots pine forest in Germany was significantly reduced when θ dropped below 0.16 m3 m-3, we did not detect any considerable effect of shallow soil drought on daily and total forest water use (Ttot = TC + ETU). Even during a 90-day period in spring 2009 (DOY 95-185) when W was continuously below 12 mm (≈ θ of 0.12 m3 m-3 - Fig. 2), we hardly detected any significant effect on daily and total Ttot (Fig. 4 and Fig. 5).
Our observed similar linear increase of Ttot with increasing GR (Fig. 6) under conditions of limiting and non-limiting shallow soil water availability reflects the energy demand for the evaporation of water ([4]) and indicates that limitations of transpiration by stomatal closure; due to low soil water availability and/or dry air played a minor role at this site ([37], [31]). This is corroborated by the dependency of daily Ttot on the vapour pressure deficit whose slope tended to decrease only at very high VPD values (Fig. 6), as also observed by eddy covariance measurements of water vapour exchange over forests at other study sites associated in the FLUXNET project ([19]).
Our estimated growing season ET averaged 291 ± 27 mm (Tab. 1) and was composed of the following fluxes: 33 ± 7% from TC, 40 ± 4% from ETU, 27 ± 5% from I (Tab. 1). These values obtained for each layer (evapo)-transpiration (TC, ETU and I - Tab. 1) are in good agreement with TC, ETU and I values reported for other P. sylvestris forest ecosystems in Europe ([36], [22], [21], [34], [12], [15]), and Central Siberia ([17], [39])
Our estimated growing season Ttot of 200 to 244 mm was within the lower end of values reported for other coniferous forest ecosystems (e.g., 90-700 mm - [19], [23], [8], [34], [12], [15], [30]). Obviously, such low total growing season Ttot values as obtained at our study site are restricted to extreme climatic and/or soil conditions ([24], [26]), and a sparse understory. In addition, the low LAI of 0.55 and the low canopy coverage of 33% ([25]), may have also contributed to the low TC (Tab. 1) and hence also to the low Ttot , although maximum whole tree water use rates obtained for P. sylvestris at our study site (25.4-8.1 kg day-1 - [37]) were in good agreement with data reported for other conifer tree species comparable in DBH (18-32 cm - [38]).
Conclusions
Precipitation measurements, scaled sap flow rates, estimates of ETU and I by the soil water budget method ([18]), and derived from the literature ([3]), respectively made it possible to investigate the water balance of an inner Alpine P. sylvestris forest which had no access to groundwater. Our data suggest that P. sylvestris forests in inner Alpine dry valleys are able to cope with high evaporative demand even under limited soil water availability (SWA) in the topsoil. In all the growing seasons investigated the amount of precipitation exceeded the total actual evapotranspiration (= TC + ETU + I). Although shallow soil drought did not play a major role at our study site, further research has to include estimates of δ18O isotope ratios of precipitation, plant tissues (xylem and foliage), and soil water in different soil depths ([29]), as well as the quantification of rooting depths ([35]). All these assessments should be combined with an evaluation of absorptive root areas in different soil layers by earth impedance methods ([6]), in order to assess the accessibility of different soil water horizons as sources for transpiration ([20]) of trees and understory vegetation in inner Alpine dry valleys and other drought-prone areas indicated by low LAI, a low canopy coverage and a sparse understory.
List of abbreviations
The following abbreviations have been used throughout the paper:
- As-stand: cumulative sapwood area per ground surface area;
- DBH: diameter at breast height;
- ET: growing season cumulative evapotranspiration;
- ETU: soil evaporation and understory transpiration;
- I: rainfall interception;
- P: precipitation;
- PET: potential evapotranspiration (Turc equation);
- Qs: sap flow density;
- GR: global radiation;
- R: total run off = RS + RD;
- RD: deep seepage;
- REW: relative extractable water;
- RH: relative humidity;
- RS: surface run off;
- SWA: soil water availability;
- T: air temperature;
- TC: canopy transpiration;
- Ttot: total forest water use (TC + ETU);
- VPD: vapour pressure deficit;
- W: shallow soil water content;
- ΔW: change in the soil water content;
θ: soil moisture.
Acknowledgements
Part of this study was financially supported by the National Austrian Science Fund Project FWF P 22206-B16 “Transpiration of conifers in contrasting environments” and FWF P25643-B16 “Carbon allocation and growth of Scots pine”. Special thanks to two anonymous reviewers for their valuable comments on the manuscript.
References
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Department of Alpine Timberline Ecophysiology, Federal Research and Training Centre for Forests, Natural Hazards and Landscape (BFW), Rennweg 1, A-6020 Innsbruck (Austria)
Walter Oberhuber
Department of Botany, Leopold-Franzens-Universität Innsbruck, Sternwartestraße15, A-6020 Innsbruck (Austria)
Corresponding author
Paper Info
Citation
Wieser G, Gruber A, Oberhuber W (2018). Growing season water balance of an inner alpine Scots pine (Pinus sylvestris L.) forest. iForest 11: 469-475. - doi: 10.3832/ifor2626-011
Academic Editor
Emanuele Lingua
Paper history
Received: Sep 06, 2017
Accepted: May 07, 2018
First online: Jul 02, 2018
Publication Date: Aug 31, 2018
Publication Time: 1.87 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2018
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 49904
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 41740
Abstract Page Views: 3975
PDF Downloads: 3132
Citation/Reference Downloads: 9
XML Downloads: 1048
Web Metrics
Days since publication: 2932
Overall contacts: 49904
Avg. contacts per week: 119.14
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2018): 7
Average cites per year: 0.88
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
The impact of land use on future water balance - A simple approach for analysing climate change effects
vol. 14, pp. 175-185 (online: 13 April 2021)
Research Articles
Coping with spring frost-effects on polyamine metabolism of Scots pine seedlings
vol. 10, pp. 227-236 (online: 27 January 2017)
Review Papers
Drought-induced mortality of Scots pines at the southern limits of its distribution in Europe: causes and consequences
vol. 3, pp. 95-97 (online: 15 July 2010)
Research Articles
Sap flow, leaf-level gas exchange and spectral responses to drought in Pinus sylvestris, Pinus pinea and Pinus halepensis
vol. 10, pp. 204-214 (online: 01 November 2016)
Research Articles
Links between phenology and ecophysiology in a European beech forest
vol. 8, pp. 438-447 (online: 15 December 2014)
Research Articles
Comparative water balance study of forest and fallow plots
vol. 5, pp. 188-196 (online: 02 August 2012)
Short Communications
Preliminary indications for diverging heat and drought sensitivities in Norway spruce and Scots pine in Central Europe
vol. 13, pp. 89-91 (online: 01 March 2020)
Research Articles
Web GIS-based simulation of water fluxes in the Miyun catchment area
vol. 7, pp. 363-371 (online: 19 May 2014)
Research Articles
Carbon and water vapor balance in a subtropical pine plantation
vol. 9, pp. 736-742 (online: 25 May 2016)
Research Articles
Slope shape effect on runoff and soil erosion under natural rainfall conditions
vol. 7, pp. 110-114 (online: 18 December 2013)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







