Fragmentation of Araucaria araucana forests in Chile: quantification and correlation with structural variables
iForest - Biogeosciences and Forestry, Volume 9, Issue 2, Pages 244-252 (2015)
doi: https://doi.org/10.3832/ifor1399-008
Published: Aug 28, 2015 - Copyright © 2015 SISEF
Research Articles
Abstract
Landscape fragmentation is one of the main threats to South American temperate forests due to population growth, conversion of native forests to plantations of exotic species and non-sustainable timber harvesting. The lack of forest connectivity can interfere with pollination, seed dispersal, biodiversity and landscape quality. Species with relatively limited seed dispersal are potentially more sensitive to the landscape fragmentation. Araucaria araucana (Mol.) K. Koch is a long-lived, slow-growing, relict conifer in South America’s temperate forests with large seeds possessing a limited dispersal range. The objective of the study was to identify priority areas for Araucaria conservation based on fragmentation quantification and correlation with structural variables and regeneration conditions. Results from the FRAGSTATS® and CONEFOR® software indicated that Araucaria connectivity has increased in sites located in the central Andean Range in comparison to other sites, because of reduced human and livestock pressure as well as the relative absence of commercial plantations. The proximity index ranged from 6.01 m to 34834.2 m, and the probability of connectivity has significantly increased (175663 ha) in the central Andean Range. Significant relationships were found between the Simpson’s index (or the probability of connectivity) and basal area, and between the mean largest patch index and crown diameter. The largest patch index (r = 0.6; p < 0.05) and the area-weighted mean proximity index (r = 0.767; p < 0.05) were the most important landscape metrics influencing Araucaria regeneration. Furthermore, the integration of spatial pattern analysis obtained from satellite images and aerial photographs with forest and regeneration characterization from field sampling allowed to identify the most vulnerable areas. The methodology presented here can assist in the identification of target areas for spatial conservation, including management needs under the current budget restrictions.
Keywords
Landscape Conservation, Landscape Metrics, Landscape Connectivity, Spatial Pattern Indicators
Introduction
Throughout the world, the commercial expansion of plantations of exotic species has led to deforestation and native landscape fragmentation ([62], [44]). Forest fragmentation is known to impact landscape quality, biodiversity and wildlife survival ([3], [48], [56], [49], [22], [6], [61]). Approximately 40% of native forests ([12]) have been destroyed due to human population growth, conversion of natural landscapes to urban use ([35]), conversion of native forests to plantations of exotic species ([45]), and non-sustainable timber harvesting ([13]).
Araucaria araucana (Mol.) K. Koch (henceforth: Araucaria) is a long-lived, slow-growing, relict conifer in South America’s temperate forests with high cultural, social and economic importance ([1], [2], [36], [32], [33]). Araucaria is listed as “vulnerable” ([34]) and a “national monument of Chile” (Supreme Decree 43 and Native Forest Law); designations that consolidate both landscape protections and harvesting prohibition for the species. Since it is illegal to cut Araucaria, there have been a number of efforts (mainly from private landowners who cover about 52% of its spatial distribution) to find benefits or profits that can be derived from these forests such as carbon storage and recreational activities.
Araucaria has two regeneration strategies: vegetative regeneration by sprouting and root suckering and germinative regeneration ([58], [23]). Araucaria seedlings tend to establish beneath parent trees because of the limited dispersal range of its large seeds ([10], [50]). If seedlings immediately colonize gaps and open areas created by dead trees, they can develop into the canopy layer ([29]). It also colonizes disturbed areas such as volcanic ash deposits and lava flows and survives in areas with suppressed conditions and wildfire ([8], [60], [27], [28]). Dynamic succession can be limited by masting, seed predation and understory vegetation effects ([50]). Although the species possesses a cone production masting pattern, environmentally-triggered, intermittent, moderately-fluctuating, and regionally-synchronous seed predation is compatible even with a low production of seeds ([26], [59]). Araucaria regeneration can be strongly altered by livestock and wildlife, mainly Sus scrofa L. ([29], [51], [63]).
High rates of deforestation and habitat fragmentation were found on the Chilean Coastal Range ([9], [14], [30]). Araucaria forests located in the Coastal Range were often surrounded by plantations due to the progressive conversion of native forests to plantations of exotic species ([9]). While plantations and urban areas are generally aggregated, native forests are often more fragmented. New business projects associated with the timber industry have expanded rapidly, requiring the development of new plantations of exotic species, mainly in areas with high site index and easy accessibility ([5]). In this sense, Araucaria forests can be classified as vulnerable because of their lack of commercial value on private lands, as well as their slow growth rate and poor regeneration in relation to vegetation competition and human disturbances ([16], [17]).
Araucaria fragmentation and the subsequent isolation of habitat patches can interfere with pollination, seed dispersal, natural regeneration and gene flow, among other ecological processes ([18], [40]). A large number of indexes have been developed to assess the ecological impacts of landscape pattern change ([31], [7]). Some studies have shown that the practical outcomes resulting from the application of some network indexes to ecological problems are very similar to those provided by considerably more complex, data-intensive, and spatially-explicit population models ([53]).
In this paper, the landscape is represented as a set of nodes (Araucaria patches) and links or connections. Landscape connectivity is analyzed within the concept of habitat availability at the landscape scale through the integral index of connectivity (IIC) and the probability of connectivity (PC - [52]). Araucaria availability is based on the comparison of the area existing within Araucaria patches (intrapatch connectivity) and the area made available through connections with other Araucaria patches in the landscape (interpatch connectivity). Recent studies have presented analysis of the equivalent connectivity (EC) for these two indices, EC(IIC) and EC(PC), to more adequately quantify landscape connectivity ([53]).
This research hypothesized that structural forest variables (Araucaria density, proportion of female trees, diameter at breast height, basal area, crown radius, canopy cover and understory cover) are correlated to the fragmentation of Araucaria patches based on landscape metrics (Simpson’s index, patch density, largest patch index, proximity index and the probability of connectivity). The overall purpose of this study was to identify structural variables and regeneration conditions (in terms of Araucaria seedlings number) and their correlation with Araucaria fragmentation using remote sensing, Geographic Information Systems (GIS), FRAGSTATS® and CONEFOR® software and field data collection. The ecological, cultural and socio-economic importance of the species requires objective tools for resource allocation under budget constraints, and this study will provide information that will help in identifying priority areas for conservation. The temporal evaluation of forest change and regeneration conditions based on permanent sampling units linked to fragmentation analysis constitutes an objective framework for assessing the degree of threat to natural ecosystems.
Methods
Study area
Chile contains more than half of the worldwide range of Araucaria, covering approximately 260 000 ha ([12]) in two separate areas: the Andean Range and the Chilean Coastal Range (Fig. 1). About 97% of the Araucaria forests are restricted to the upper elevations of the Andean mountain range from Region VIII to Region XIV.
Fig. 1 - Location of the study sites and A. araucana forest range in central Chile.
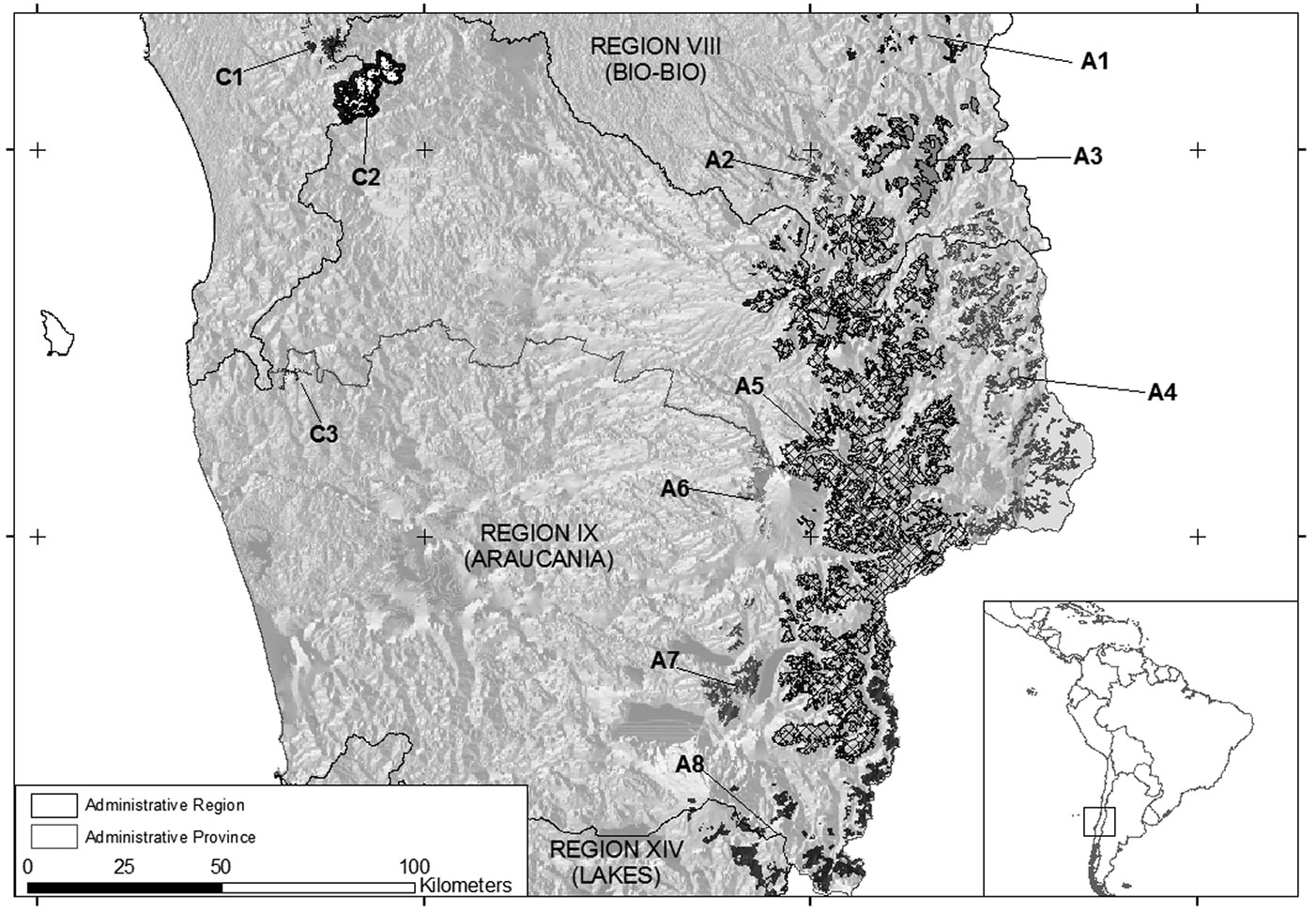
The species is located in a wide altitudinal and ecological range. In the Andean Range most of the soils are derived from ash deposited by volcanic activity ([43]); however, the Chilean Coastal Range possesses better developed soils (granite and metamorphic textures - [47]). The climate of both mountain ranges has a Mediterranean influence reflected by a winter-maximum in precipitation and relatively dry summers ([4]). Annual precipitation varies between 1500 and 3000 mm, although at higher altitudes the precipitation can reach more than 4000 mm, the majority falling as snow. Mean annual temperature is highly variable with minimum temperatures in July and maximum temperatures in February.
Spatial configuration of Araucaria forests
We used ArcGIS® 9.3 ([25]) and ERDAS® 9.3.2 ([24]) to analyze the conservation and fragmentation condition of Araucaria ecosystems in Chile. Firstly, we identified Araucaria distribution using the National Mapping of Native Vegetation Resources ([12]). In order to get a finer spatial resolution for its current distribution (30 m), a set of Landsat scenes (Thematic Mapper and Enhanced Thematic Mapper) were mosaicked to be used as background for mapping (a list of cartographic materials used is in Tab. 1). Because of the size of the mosaic, a raster catalog of mosaicked map for each Araucaria site was used to organize all sites into a single countrywide database using ArcGIS. Aerial photographs were acquired because they presented advantages over remote sensing images in reference to Araucaria crown identification. We developed a single Araucaria map based on these cartographic materials (spanning from 2006 to 2011). The Delphi method was used to delimit homogeneous Araucaria sites according to geographic (longitude, latitude, accessibility and human pressures), climatologic (temperature and precipitation) and ecological (canopy and understory composition) attributes along the species distribution. Delphi is an expert survey consisting of two or more “rounds”, in which previous rounds are given as feedback in later rounds ([39]). Experts participated in a later round and agreed on the identification of eleven Araucaria sites (Tab. 2 and Tab. 3).
Tab. 1 - Cartographic material used to Araucaria forests delimitation.
| Cartographic material |
Scale or spatial resolution (m) | Year |
|---|---|---|
| National Mapping of Native Vegetation | 1:50000 | 1999 |
| Aerial photographs (National Forest Corporation) | 1:100000 | 2006-2007 |
| Aerial photographs (private landowners) | 1:50000, 1:20000 | 2009-2010 |
| Aerial photographs (Forestry companies) | 1:20000, 1:10000 | 2010-2011 |
| Landsat scenes (Thematic Mapper) | 30 | 1/1/2011, 2/2/2011 |
| Landsat scenes (Enhanced Thematic Mapper) | 30 | 6/1/2010, 8/12/2010 |
| Google Earth | dependent on the site | 2011 |
Tab. 2 - Andean Range sites. (Tm): mean temperature range: (P): annual precipitation.
| Site | Location | Description | Climate | Vegetation | Observations |
|---|---|---|---|---|---|
| A1 | Northern border of Andean Range | It is a discontinuous site where A. araucana fragments are variable and found predominantly at altitudes higher than 1500 m in areas with moderate slopes. Its climate is very similar to the Coastal Range | Tm: 5.7-24 °C P: 1800-2050 mm |
N. dombeyi, Griseliniascandens (Ruiz et Pav.) Taub, Desfontainea spinosa (Ruiz et Pav.), C. culeou (E. Desv), ... | Private land. A. araucana is generally in old grown stage |
| A2 | Northern Andean Range | This small site has a great competence to seedling establishment because of the dense understory presence. The climate shows a dry period in the summer months (less than four months). A. araucana found predominantly at altitudes lower than 1.500 m | Tm: 5.7-24 °C P: 1800-2700 mm |
Chusquea culeou, herbaceous species and Nothofagus antarctica (G.Forster) Oerst, ... | Private land. New hydrological project impact |
| A3 | North-eastern Andean Range | It is a discontinuous site located in Region VIII, near the outer edge of Araucaria distribution. This site is similar to “Area A2” but it is at higher altitudes | Tm: 5-20 °C P: 1900-2800 mm |
Nothofagus pumilio (P. et E.) Krasser (at medium altitudes) and N.antarctica, ... | Steep topography (high slopes). Poor accessibility |
| A4 | Eastern Andean Range | This site is on the border of Argentina. The area is located at higher altitude, and most of the precipitation falls as snow, covering soils throughout the year. The climate is cold with temperatures less than 0°C during most months of the year | Tm: -2.5-14.5°C P: 3000-3800 mm |
N. antarctica, ... | Perpetual snow. Vegetation is limited because of climate conditions |
| A5 | Central Andean Range | This is the main zone of Araucaria distribution in Chile showing different natural protected areas. The topographic and climate conditions depend on altitude and aspect | Tm: 0.5-16 °C P: 2000-3500 mm |
N. dombeyi (Mirb.) Oerst (at lower altitudes), N. pumilio (at middle altitudes) and N.antarctica (at higher altitudes), C.culeou, Drimys winteri (J.R. et Pav.) Taub, Podocarpus saligna (D.Don), Austrocedrus chilensis (D.Don) Pic.Sem. et Biz., ... | Recent large wildfires. Perpetual snow. Volcanic ash deposition in lava flows |
| A6 | Western Andean Range | This area has lower altitudes than other Andean sites because of its position in the pre-Andean Range. Climate is moist temperate | Tm: 2-23 °C P: 1500-2500 mm |
N. dombeyi, C.culeou, Persea lingue (Ruiz et Pav.), Berberis spp., Ribes spp.,... | A. araucana is generally surrounded by native forests (Nothofagus spp.). Volcanic soil. Cattle |
| A7 | South-western Andean Range | It is located in the pre-Andean Range in a discontinuous shape of the main site (Area A5). The area has a worse accessibility than adjacent sites (“Area A5” and “Area A6”) | Tm: 6-15.1 °C P: 1500-2500 mm |
Araucaria-Nothofagus forests, mainly N. dombeyi, C.culeou, Berberis pearcei (Lecchler.),... | Agricultural and grasslands presence. Firewood exploitation. Volcanic soil |
| A8 | Southern Andean Range | The most southern site of the Chilean distribution and the bordering stands of Argentina. Forests are mixed stands based on the altitude and aspect. Generally, Araucaria-Nothofagus forests has a dense understory | Tm: 2.1-21.4 °C P: 2000-3500 mm |
N. dombeyi and N. pumilio (at medium altitudes), N.antarctica (volcanic soils in upper areas), C.culeou, B.pearcei, Chiliotrichium rosmarinifolium L., ... | Most of the soils are derived from ash deposited by volcanic activity. Protected and recreational area |
Tab. 3 - Chilean Coastal Range sites. (Tm): mean temperature range: (P): annual precipitation.
| Site | Location | Description | Climate | Vegetation | Observations |
|---|---|---|---|---|---|
| C1 | Northern Chilean Coastal Range | Topography is characterized as wavy. Two dwarf populations of A. araucana were found at the top of this area | Tm: 6.9-20 °C P: 1.500-2.000 mm |
N. dombeyi, Griselinia scandens, Desfontainea spinosa, C. culeou, ... | Private land. Pressure of exotic species (plantations) |
| C2 | Nahuelbuta National Park | This area shows the highest altitudes of the Coastal Range. In this area, precipitation could reach 3.000 mm with a dry month | Tm: 5.9-19.2 °C P: 2.000-3.000 mm |
Nothofagus spp. (dependent on the altitude gradient), C. culeou, Drimys winteri, Azara lanceolata (Hook F.), Valeriana lapathifolia (Vahl.), ... | Protected and recreational area. Rocky surface. Recent surface wildfires |
| C3 | Southern Chilean Coastal Range | It is a remnant site, known as “La Cabaña”, located at lower elevation (about 600 m). Mean temperature drops in relation to other sites of the Coastal Range | Tm: 9-15.7 °C P: 1.500-2.500 mm |
It is frequently accompanied by evergreen species | Private land. Pressure of exotic species (plantations) |
Satellite images were corrected geometrically, atmospherically and topographically using ERDAS. Preparing the images required the transformation to the WGS84 Zone 19S system, creating an overlap of the Araucaria site images in vector format using ArcGIS. The scale used allowed the identification of the following land cover types: native forest without presence of Araucaria (NF), native forest with presence of Araucaria (NFA), shrubland (S), plantations of exotic species (P), agricultural and pasture lands (A), water and wetlands (W), urban areas (U) and bareland (B). Pixels were assigned to the land cover type of highest probability based on a supervised classification of overlapping signatures and the maximum likelihood criterion ([11]). Accuracy assessment involved identifying a set of sample locations (control points) that were sampled in the different field trips. A confusion matrix was obtained to compare the class identified for each control point with the land cover derived from the satellite images and aerial photographs. Overall accuracy and the Kappa index were calculated for each land cover type. Accuracy assessment was validated using a dataset of 313 random control points covering eleven Araucaria sites: eight sites in the Andean Range (Tab. 2) and three sites in the Chilean Coastal Range (Tab. 3). Then, post-classification modifications were mapped using cartographic information in GIS.
Scale refers to the “grain” or the spatial resolution of the map and “extent” refers to the map size ([57], [37]). Fine spatial resolution can identify Araucaria patches within native forest patches using aerial photographs from public agencies, forestry companies and private landowners. The boundaries of Araucaria forests were identified based on the forests’ crown architecture, although there were some difficulties in pixels where Araucaria was not reaching the upper canopy layer. A minimum mapping unit of more than 0.45 ha was used (similar to other studies) because of the difficulty of small fragment assessment ([20]). Although the validation of Araucaria patches was done in the field, satellite images and aerial photographs were the only suitable tool for the geographic scale used in assessing various remote areas.
Additional information was collected by field trips in five administrative provinces: Arauco and Bio-Bio in Region VIII, Cautin and Malleco in Region IX and Valdivia in Region XIV. These provinces cover twenty municipalities (the administrative division of each province). Field trips were largely random with the exception of the edges of Araucaria sites, generally the most vulnerable areas to landscape fragmentation ([30]).
Spatial pattern analysis
The description of spatial patterns requires information regarding the composition (or variability) and configuration (or arrangement) of the Araucaria patches. Some landscape metrics were selected based on principles of forest fragmentation and the general guidelines set by various authors ([31], [7]). We identified six landscape metrics: the Simpson’s diversity index (SI), patch size (PS), patch density (PD), largest patch index (LPI), radius of gyration (GYRATE) and proximity index (PROX) by FRAGSTATs software ([41]). Simpson’s index was calculated as one minus the sum of the proportional areas of each patch type squared. We calculated the Simpson’s index both as a diversity of patch sizes and a diversity of land use types: NF, NFA, S, P, A, W, U and B using remote sensing images. Patch density (number of patches per 100 ha) cannot be independently interpreted as a measure of patch size. Area-weighted mean patch size was used to reduce problems related to skewed distributions of patch sizes inside each site. Largest patch index quantifies the percentage of total landscape area comprised by the largest Araucaria patch. Area-weighted mean radius of gyration (average extensiveness in meters of connected cells) is a measure of both fragment extent and habitat connectivity ([20]). Area-weighted mean proximity index (ratio between the area and the distance of patches) measures the degree of fragmentation of the Araucaria patches under three scenarios or search buffers: 250, 500 and 1000 m, based on the size of the different sites. When the search buffer extended beyond the landscape boundary, only patches within the landscape were considered in the computations. Proximity index increases as the neighborhood (defined by the search radius) is increasingly occupied by patches of the same class, and it increases as those patches become closer and more contiguous. We constructed a correlation matrix for these landscape metrics (Tab. 4) and discarded some that were closely correlated with others (r > 0.7). The Simpson’s index based on patch size, patch size and radius of gyration were discarded because of their correlations with patch density and proximity index.
Tab. 4 - Matrix of the Spearman’s correlation coefficients of landscape metrics for the different study sites. (SI): Simpson’s index calculated as (1) diversity of patch sizes and (2) as diversity of forest types; (PS): patch size; (PD): patch density, (LPI): largest patch index; (GYRATE): radius of gyration; (PROX): proximity index; EC(ICC), EC(PC): see definition in the text; (*): P < 0.05; (**): P < 0.01.
| Metrics | SI1 | SI2 | PS | PD | LPI | GYRATE | PROX | EC(ICC) | EC(PC) |
|---|---|---|---|---|---|---|---|---|---|
| SI1 | 1 | - | - | - | - | - | - | - | - |
| SI2 | 0.082 | 1 | - | - | - | - | - | - | - |
| PS | 0.6 | 0.036 | 1 | - | - | - | - | - | - |
| PD | 0.764** | 0.055 | 0.50 | 1 | - | - | - | - | - |
| LPI | 0.564 | 0.155 | 0.44 | 0.52 | 1 | - | - | - | - |
| GYRATE | 0.764** | 0.291 | 0.72* | 0.54 | 0.773** | 1 | - | - | - |
| PROX | 0.68* | 0.091 | 0.62* | 0.42 | 0.64* | 0.92** | 1 | - | - |
| EC(ICC) | - | 0.264 | - | -0.409 | 0.091 | - | 0.382 | 1 | - |
| EC(PC) | - | 0.136 | - | -0.409 | -0.009 | - | 0.318 | 0.982** | 1 |
Then, we used the CONEFOR SENSINODE software ([52]), which can characterize the degree of habitat connectivity for each Araucaria sites (mosaic of Araucaria patches), providing an idea of the current status of the Araucaria landscape. EC(IIC) and EC(PC) are defined as the size of a single habitat patch (maximally connected) that would provide the same value of the IIC and PC metric as the actual habitat pattern in each site. EC(IIC) and EC(PC) was computed as the square root of the numerator of the IIC and PC indices (Appendix 1). We only used EC(PC) and discarded EC(ICC) that was closely correlated (Tab. 4). EC(PC) index presents the advantages of having area units, a more reasonable and usable range of variation, and, more importantly, an easier and straightforward interpretation especially when directly compared with the temporal changes in habitat area ([53]).
Kruskal-Wallis non-parametric test was used to compare eleven Araucaria sites that are independent and that have different sample sizes. The parametric equivalent of the Kruskal-Wallis test is the one-way analysis of variance (ANOVA). Dunn’s non-parametric post-hoc test helped identify the significant relationships among Araucaria sites and the number of Araucaria patches (χ2 = 18.99, p = 0.04), Simpson index based on land uses types (χ2 = 18.88, P = 0.045), patch density (χ2 = 19.63, p = 0.03), largest patch index (χ2 = 20.09, p = 0.028), proximity index (χ2 = 31.95, p = 0.001) and ECC(PC) (χ2 = 40.54, p = 0.01).
Structural variables and regeneration in Araucaria forests
The forest inventory was carried out in circular plots of 1000 m2 using a stratified random sampling method. Different sampling plots were established according to Araucaria density identified by images and photographs within each location. The number of plots for each site was established proportional to the Araucaria area with the exception of the C3 site, which was not considered. The sample amounted to 104 plots (4 in A1, 5 in A2, 8 in A3, 12 in A4, 45 in A5, 5 in A6, 5 in A7, 11 in A8, 5 in C1, 4 in C2) located across the different Araucaria patches, incorporating variables such as stand density (trees ha-1), Araucaria density (trees ha-1), proportion of female trees (%), mean diameter at breast height (cm), basal area (m2 ha-1), mean crown radius (m), canopy cover (%) and understory cover (%). We assessed the relationship between Simpson index according to land use types, patch density, largest patch index, proximity index, EC(PC) and these structural variables. A Spearman’s correlation test (non-parametric relationship) was used to identify landscape metrics that had significant relationships with forest structural variables.
Sampling data were collected for both forest structural variables and regeneration parameters in order to assess landscape condition. Regeneration data were collected in 974 subplots (within 1000 m2 plots) containing Araucaria and seedlings of other species. At least six subplots (random 100 m2 plots) were located within each sample unit. This plot size had some limitations because of the presence of “seed trees” near the plot borders. Consequently, some Araucaria seedlings were either present inside or outside of the plot. Regeneration was expressed as the number of seedlings per hectare to allow the model to be applied independently of plot size. Although the regeneration behavior for shade-tolerant species was consistent with the regeneration hypothesis associated with the competition-colonization model and spatial patterns ([16]), seedlings had higher probability of reaching the canopy height in gaps created by dead trees. In this sense, all seedlings were classified by three height classes (< 50 cm, 50-130 cm and > 130 cm) according to other regeneration studies ([19]). When Araucaria regeneration is less than 50 cm, seedlings are exposed to competition with shrub species, mainly Drimys winteri var. andina and Desfontainia spinosa, and Nothofagus, which have seedlings that colonize gaps faster than Araucaria. Seedlings over 130 cm in height can reach a further crown developing or polewood stage (when the tree occupies the dominant layer in the canopy - [19]). In addition to the landscape fragmentation analysis, a Spearman’s correlation test was used to determine if there were significant relationships (r > 0.6) between landscape metrics and Araucaria regeneration.
Results
Spatial configuration of Araucaria forests
Classification accuracy was 86.21% based on overall agreements (Tab. 5). The lowest values of user accuracy were obtained for native forests (78.13%) and shrublands (78.57%). Native forest (NF) was confused with water and wetland (W), likely due to similarly dark spectral responses. We used a false color image from the satellite images to solve this misclassification problem. The false-color image showed the scene in red. While plants reflected more infrared so they were brighter red, water was black and blue. Shrubland (S) was more frequently miss-reported as agricultural and pasture land, also likely due to their similar spectral responses. Land cover cartography based on satellite images was improved using aerial photography (focusing on Araucaria crown architecture) and field trips. Control points were important for distinguishing between native forest without/with the presence of Araucaria. We analyzed Araucaria sites (Tab. 2, Tab. 3) to calculate the area and percentage of each land cover class (Tab. 6, Tab. 7). “Area C3” showed the smallest Araucaria area due to the expansion of exotic species plantations (64.65% of the total area). Sites located on the Andean Range had less pressure from plantation establishment due to steep topography and difficult accessibility (ranging from 0% to 3.53% of the total area). Andean fragments were configured as a landscape mosaic with a very large patch (A5) surrounded by smaller sites. Shrubland was frequently located within the limits of Araucaria distribution, such as A4 and A1 sites (23.07% and 19.75%, respectively). Bareland was commonly found in some Andean sites such as A4, A5 and A8. While A4 was located in relation to perpetual snow, A8 was related to volcanic activity, and A5 contained both perpetual snow and volcanic activity. Field trips showed other perturbations such as forest fires in A5 and C2, but Araucaria trees survived after wildfire due to its bark thickness. A new hydrological project, located between A2 and A5, proved an important anthropogenic perturbation due to the increase in the density network and land use changes.
Tab. 5 - Confusion matrix based on satellite images and aerial photographs. (NF): native forest; (S): shrubland; (P): plantation of exotic species; (A): agricultural and pasture lands; (W): water and wetlands; (U): urban area; (B): bareland.
| Classified information |
Reference data | User’s accuracy (%) |
Kappa index |
||||||
|---|---|---|---|---|---|---|---|---|---|
| NF | S | P | A | W | U | B | |||
| NF | 50 | 6 | 0 | 1 | 7 | 0 | 0 | 78.13 | 0.7518 |
| S | 0 | 44 | 2 | 9 | 0 | 1 | 0 | 78.57 | 0.5956 |
| P | 1 | 0 | 37 | 1 | 0 | 0 | 0 | 94.87 | 0.9358 |
| A | 2 | 0 | 1 | 50 | 0 | 1 | 0 | 92.59 | 0.9133 |
| W | 0 | 0 | 0 | 5 | 33 | 0 | 0 | 86.84 | 0.8421 |
| U | 0 | 0 | 0 | 0 | 0 | 34 | 0 | 100 | 1 |
| B | 0 | 0 | 0 | 0 | 2 | 0 | 26 | 92.86 | 0.9205 |
| Producer’s accuracy (%) |
94.34 | 88 | 92.5 | 75.76 | 78.57 | 94.44 | 100 | - | - |
| Overall classification accuracy | 86.21 | - | |||||||
| Total Kappa Index | - | 0.8407 | |||||||
Tab. 6 - Area of each land cover class (in ha) at the A. araucana sites using a 1000 m buffer. (NF): native forest without presence of A. araucana; (NFA): native forest within presence of A. araucana; (S): shrubland; (P): plantations of exotic species; (A): agricultural and pasture lands; (W): water and wetlands; (U): urban area; (B): bareland.
| Land use | A1 | A2 | A3 | A4 | A5 | A6 | A7 | A8 | C1 | C2 | C3 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| NF | 13365 | 16201.49 | 22931.11 | 32785.15 | 150954.48 | 7082.09 | 14686.87 | 27876.54 | 9867.12 | 9661.36 | 693.83 |
| NFA | 3475.58 | 3352.35 | 14245.27 | 23269.22 | 161512.22 | 4947.9 | 8191.02 | 25082.33 | 2992.71 | 6336.7 | 691.18 |
| S | 5410.71 | 2407.72 | 7049.01 | 31015.86 | 54022.77 | 2731.28 | 1917.31 | 5089.1 | 344.95 | 436.62 | 133.98 |
| P | 83.33 | 596.09 | 4.8 | 0 | 826.18 | 642.37 | 443.29 | 339.9 | 1194.9 | 1402.1 | 3626.8 |
| A | 3021.79 | 154.73 | 4621.46 | 25503.39 | 26310.91 | 457.62 | 1059.22 | 4830.55 | - | 723.01 | 41.82 |
| W | 40.34 | 86.99 | 277.49 | 116.69 | 2889.68 | 192.81 | 648.52 | 379.17 | 0 | 0 | 0 |
| U/B | 2049.57 | 24.24 | 6753.01 | 21771.41 | 49528.93 | 2137.37 | 68.58 | 9583.02 | 0 | 166.05 | 422.18 |
| Total | 27446.33 | 22823.62 | 55882.2 | 134461.72 | 446045.18 | 18191.45 | 27014.82 | 73180.62 | 14399.71 | 18725.84 | 5609.79 |
Tab. 7 - Percentage of each land cover class (%) at the A. araucana sites using a 1000 m buffer. (NF): native forest without presence of A. araucana; (NFA): native forest within presence of A. araucana; (S): shrubland; (P): plantations of exotic species; (A): agricultural and pasture lands; (W): water and wetlands; (U): urban area; (B): bareland.
| Land use | A1 | A2 | A3 | A4 | A5 | A6 | A7 | A8 | C1 | C2 | C3 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| NF | 48.70 | 70.99 | 41.03 | 24.38 | 33.84 | 38.93 | 54.37 | 38.09 | 68.52 | 51.59 | 12.37 |
| NFA | 12.66 | 14.69 | 25.49 | 17.31 | 36.21 | 27.20 | 30.32 | 34.27 | 20.78 | 33.84 | 12.32 |
| S | 19.71 | 10.55 | 12.61 | 23.07 | 12.11 | 15.01 | 7.10 | 6.95 | 2.40 | 2.33 | 2.39 |
| P | 0.30 | 2.61 | 0.01 | 0 | 0.19 | 3.53 | 1.64 | 0.46 | 8.30 | 7.49 | 64.65 |
| A | 11.01 | 0.68 | 8.27 | 18.97 | 5.90 | 2.52 | 3.92 | 6.60 | 0 | 3.86 | 0.75 |
| W | 0.15 | 0.38 | 0.50 | 0.09 | 0.65 | 1.06 | 2.40 | 0.52 | 0 | 0 | 0 |
| U/B | 7.47 | 0.11 | 12.08 | 16.19 | 11.16 | 11.75 | 0.25 | 13.10 | 0 | 0.89 | 7.53 |
Spatial pattern analysis
Landscape fragmentation analysis requires to consider both the patch size and the spatial configuration of Araucaria patches. While patch size was analyzed based on the number of patches, patch density and the largest patch index (the patch size was discarded previously), the quantification of the Araucaria patch connectivity was conducted according to the mean proximity index (PROX) and EC(PC). Significant relationships were found between Araucaria sites and landscape metrics. An analysis of the number of patches differences was performed among Araucaria sites showing six significant groups (Tab. 8). For example, in the Coastal Range differences were dtected for C1 in relation to C2 and C3. Higher values for the Simpson’s index were recorded by A3, A8, C1, C2 and C3 (“group c”) in relation to the remaining sites. Based on patch density, the highest value in C3 (“group c”) was associated with the small size of the patches. Five significant groups of Araucaria sites could be defined based on Araucaria patch density: A1, A2 and C1 (“group a”); A3, A5 and A8 (“group b”); A4 and C3 (“group c”); C2 (“group d”); and A7 (“group e”). Differences among the sites within this same group were observed in terms of the largest patch index. As an example, the largest patch index increased from 4.4% to 15.45% in the “group a” of patch density. In the Coastal Range, the largest patch index was significantly increased in C2 (“group e”) compared to C1 and C3 (“group c”).
Tab. 8 - Area-weighted landscape metrics assigned to each A. araucana site. Mean values in columns followed by the same letter are not significantly different (p > 0.05). (*): Mean values ± standard deviation obtained under three edge zones of depth: 250, 500 and 1000 m. (NP): number of patches; (SI): Simpson’s index calculated as the diversity of land uses types. For the other column labels, see Tab. 4.
| Site | NP | SI | PD (n/100 ha) |
LPI (%) |
PROX* (m) |
ECC(PC) (ha) |
|---|---|---|---|---|---|---|
| A1 | 38a | 0.52a | 0.12a | 4.4a | 6.01 ± 1.16 a | 1680.01a |
| A2 | 32a | 0.21b | 0.13a | 2.81b | 72.27 ± 10.90 b | 2919.07a |
| A3 | 34a | 0.82c | 0.054b | 12.83c | 109.40 ± 27.04 b | 11376.61ab |
| A4 | 246b | 0.50a | 0.16c | 2.03b | 142.63 ± 15.22 b | 9908.23ab |
| A5 | 339c | 0.43b | 0.067b | 20.91d | 34834.20 ± 365.49 c | 175663b |
| A6 | 10d | 0.29b | 0.045d | 30.13e | 268.08 ± 0.34 b | 5873.46ab |
| A7 | 9d | 0.36b | 0.026e | 15.38c | 986.01 ± 0 d | 8747.17ab |
| A8 | 57e | 0.73c | 0.061b | 8.51a | 230.19 ± 20.68 b | 15477.06ab |
| C1 | 17f | 0.68c | 0.1a | 15.45c | 91.63 ± 4.86 b | 2845.96a |
| C2 | 11d | 0.76c | 0.05d | 40.97e | 328.19 ± 0.02 e | 7986.07ab |
| C3 | 10d | 0.63c | 0.18c | 15.42c | 10.39 ± 0.02 a | 872.84a |
Fragmentation was accompanied by a reduction of the area-weighted mean proximity index, ranging from 6.01 m in A1 to 34834.2 m in A5 (Tab. 8). Mean proximity index identified five significant groups: A1, C3 (“group a”); A2, A3, A4, A6, A8 and C1 (“group b”); A5 (“group c”); A7 (“group d”); and C2 (“group e”). No substantial difference between mean proximity indexes calculated at different distances was recorded. According to this result, a distance of 500 m could be selected as an intermediate value between other distances. For example, in the Coastal Range the area-weighted proximity index increased in C2 in relation to C1 and C3. Proximity index pointed to the connectivity of A5, followed by A7 (values higher than 500 m). An analysis of the EC(PC) was performed among Araucaria sites showing three significant groups (Tab. 8). There was a wide difference in Araucaria connectivity ranging from 872.84 ha to 175663 ha. Higher values for the EC(PC) were recorded by A5 (“group b”) in relation to the rest of sites. Based on the probability of connectivity, the lowest value in C3, A1, C1 and A2 (“group a”) was associated with the small connectivity of the Araucaria patches.
Structural variables and regeneration condition of Araucaria forests
Although there were not enough data to support significant relationships between landscape metrics and some structural variables (such as mean diameter at breast height, canopy cover, Araucaria density and proportion of female trees), significant relationships (Spearman’s test) were found between the basal area and the Simpson’s index (calculated as a diversity of land use types) or the probability of connectivity (r = 0.92, p < 0.01 and r = 0.676, p < 0.05, respectively), as well as between the mean largest patch index and the crown diameter (r = -0.71, p < 0.05).
Regeneration condition was measured in Araucaria seedlings (mean number of seedlings per hectare) according to the sampling design for each patch. This regeneration condition was significantly correlated to the largest patch index (r = 0.6, p < 0.05) and to the area-weighted mean proximity index (r = 0.767, p < 0.05). Despite such correlation was not very high, it could explain more than 75% of the sampling units, showing an important approach to identification of potential regeneration conditions from landscape fragmentation studies. In this sense, a reduction of mean proximity index was generally related to a decline in the regeneration condition of the fragments.
Discussion
Although some authors have recommended a multi-temporal analysis of remote sensing scenes ([20], [21], [5]), we used the current spatial identification of Araucaria patches acquired in an accuracy assessment according to the confusion matrix (Tab. 5). In future studies, we will focus on Araucaria changes based on the spatial distribution achieved in this investigation. The overall accuracy of satellite imagery revealed that the supervised classification and the false-color image provided a suitable identification of land uses. Aerial photographs improved the identification of Araucaria inside of native forest land. The Andean Range has a lower level of accessibility than the Coastal Range due to the topography and the lower road network density. It was difficult to design sampling units and field trips along the entire distribution of the species, thus increasing the importance of satellite images and aerial photography to support land uses classification.
Area-weighted landscape metrics (patch size, patch density, largest patch index, Simpson’s index, radius of gyration and proximity index) and the probability of connectivity were used to analyze the spatial pattern of eleven Araucaria sites. These landscape metrics have been frequently related to forest fragmentation by scientific studies ([31], [7], [21], [38], [53]). In our study, these metrics were objective indicators in the identification of “Area C3” and “Area A1” as fragmented landscapes. Low patch connectivity was found in the Coastal Range similar to previous fragmentation approaches ([9], [20]). In this sense, the remaining patches were greatly affected by plantations of exotic species (64.65% of the total area). Sites with low Araucaria area and more fragmented spatial pattern (like in “Area C3” and “Area A1”) will have a higher probability of suffering large decreases in connectivity than other sites with large and compact patches (like in “Area A5” and “Area A8”).
The fragmentation of natural landscapes could lead to changes in vegetation composition and forest structure over the medium and long term ([30], [5], [54]). In our study, the Simpson’s index (calculated as a diversity of land uses types) and the EC(PC) significantly increased in relation to the basal area of the patches. This positive correlation could be due to the higher levels of biodiversity and basal area of the primary forests. A reduction in Simpson’s index and in the probability of connectivity could be caused by the expansion of other land uses (mainly agricultural lands and plantations of exotic species) and an increase in total edge, and as a consequence, the creation of suitable sites for shrub establishment. Araucaria path density is higher in areas altered by humans than in other areas dominated by primary or virgin forests. In this sense, basal area is indirectly related to patch size, a result that has been found in other studies ([55], [5], [21]). Conversely, largest patch index was negatively correlated to crown diameter. This relationship can be causality but it also could be related to the significant correlation between crown radius and stand density (r = -0.867, p < 0.01). The abundance of tree species increases in large patches and edge areas ([30], [21]). Stand density should decrease the mean crown radius due to the competition among canopy tree species. Then, stand density could be higher in larger patches than smaller ones because of the human disturbances and the increase in edge areas of other species.
We hypothesized that regeneration condition is correlated with the fragmentation of Araucaria patches due to human disturbances and surrounding land use changes. A reduction in patch size leads to differences in seed production ([15]) and microclimatic changes ([46]). Shade tolerant species appear to be more vulnerable to forest fragmentation than shade intolerant species because they require a certain degree of canopy protection during the first stages of growth ([9]). Site conditions in the larger forest patches facilitate the establishment and growth of these shade-tolerant species. A reduction of largest patch index was often related to a decline in the Araucaria seedlings. Larger Araucaria patches facilitated the establishment of shade-tolerant species. Therefore, the largest patch size is negatively associated to the edge length, so a reduction in patch size also leads to a dense establishment of invasive species ([30], [21], [6]). The increased density of fast-growing species may cause a decline in the abundance of Araucaria seedlings affecting its long-term survival similar to other species ([9]). Our results suggest that mean proximity index was the more accurate landscape metrics to identify Araucaria regeneration. In this sense, forest connectivity may be more important than patch size in explaining changes in woody plant composition and forest structure, similar to other approaches ([55], [42]). Species with relatively limited seed dispersal could be potentially more sensitive to changes in the landscape matrix.
Our results indicated that gradual fragmentation is associated with changes in the structural and regeneration condition of Araucaria forests. Its long-term conservation requires the improvement of the condition of some remnant fragments, according to its cultural, social and ecological value. Special attention should be given to the sites located in the Coastal and the pre-Andean Range. Indeed, these sites showed the smallest patches, and least proximity and connectivity of Araucaria patches, and as a consequence, they are the most vulnerable landscapes over the medium and long term. Likewise, molecular markers have highlighted some of these sites as priority areas of conservation because of their high levels of genetic diversity ([40]). If the fragmentation process is maintained, the regeneration may be dramatically reduced. In this sense, Araucaria sites must be monitored over time and space at the landscape level. Private lands must focus on environmental services (carbon storage) and landscape goods (leisure and recreational activities) as innovative approaches for Araucaria conservation and management. Technical recommendations of the conservation priority could be established by the integration of landscape fragmentation, forest structural variables and regeneration condition. In terms of Araucaria landscape conservation, four recommendations could be given: (1) to provide incentives for the creation of biological corridors which would increase the genetic exchange of C1 and C2 sites (indicators given by other studies based on Araucaria genetic diversity should be considered - [18], [40]); (2) to create new private protected areas and to establish collaboration agreements with private companies concerning the effects on remnant patches of A4, A5 and A8 sites, according to Araucaria fragmentation, general forest characteristics and regeneration condition; (3) to invest in tree plantation agreements with private landowners for the enrichment, densification and rejuvenation of C3, A1 and A2 stands, based on Araucaria density, seed trees presence and old-grown stage; and (4) to establish actions to conserve the Araucaria landscape based on the principles of the new Native Forest Law N° 20.283 (sites A3, A6 and A7).
Conclusions
Araucaria araucana is characterized by a wide spatial distribution in Chile. Nevertheless the Araucaria landscape is presently associated with an increase in fragmentation due to human, livestock and wildlife pressures coupled with commercial plantations, mainly in the Coastal Range. Landscape fragmentation will lead to changes in forest structure and floristic composition. The conservation approach proposed in this study is based on the landscape fragmentation, forest structural variables and regeneration condition. The identification of Araucaria fragmentation using GIS, FRAGSTATS® and CONEFOR® increases the flexibility of this methodology, enabling an extrapolation to other territories and species. Forest data used as input in our method are easy to obtain from forest inventories. We found that the Simpson’s index (calculated as a diversity of land uses types), the largest patch index, the area-weighted mean proximity index and the probability of connectivity are closely correlated with forest structural variables and the regeneration condition. Future studies should test if these correlations can be used to identify priority areas as an integration of landscape metrics and forest variables.
The great cultural, social and ecological importance of the Araucaria landscape calls for new tools, such as those presented here, which help analyze the potential effects of forest policy changes and contribute to sustainable landscape management. An adequate territorial planning could improve the effectiveness of conservation and land use allocation decisions based on the maintenance of Araucaria connectivity. The limitation of multi-temporal analysis inherent to this approach should be also considered. Further analyses should contemplate field sampling over time, constraints on the relationship between landscape fragmentation and forest structure, regeneration condition, and genetic diversity.
Acknowledgments
This research was partially supported by grants No. 207-141-018-1.0 from the Research Service of University of Concepción (Chile), grants A/023099/09 and A/030789/ 10 of the Spanish Agency of International Cooperation for the Development from the Spanish Ministry for Foreign Affairs and Cooperation. MAM is grateful to the Secretaría General de Ciencia y Tecnología de la Consejería de Economía, Competitividad e Innovación from the Regional Government of Extremadura (Spain) for the financial support.
References
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Supplementary Material
Authors’ Info
Authors’ Affiliation
Miguel Ángel Herrera
Departamento de Ingeniería Forestal, Escuela Técnica Superior de Ingeniería Agronómica y de Montes, Edificio Leonardo Da Vinci, Campus de Excelencia Internacional Agroalimentario (ceiA3), Universidad de Córdoba, ES-14071 Córdoba (Spain)
Departamento de Ingeniería del Medio Agronómico y Forestal, Avda. Virgen del Puerto 2, Centro Universitario de Plasencia, Universidad de Extremadura, ES-10600 (Spain)
Departamento de Manejo de Bosques y Medioambiente, Facultad de Ciencias Forestales, Universidad de Concepción, Casilla 160-C, Concepción (Chile)
Departamento de Genética, Escuela Técnica Superior de Ingeniería Agronómica y de Montes, Edificio Gregor Mendel, Campus de Excelencia Internacional Agroalimentario (ceiA3),Universidad de Córdoba, ES-14071 Córdoba (Spain)
Corresponding author
Paper Info
Citation
Molina JR, Martín Á, Drake F, Martín LM, Herrera MÁ (2015). Fragmentation of Araucaria araucana forests in Chile: quantification and correlation with structural variables. iForest 9: 244-252. - doi: 10.3832/ifor1399-008
Academic Editor
Matteo Garbarino
Paper history
Received: Jul 14, 2014
Accepted: May 04, 2015
First online: Aug 28, 2015
Publication Date: Apr 26, 2016
Publication Time: 3.87 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2015
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 54930
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 43993
Abstract Page Views: 4257
PDF Downloads: 5131
Citation/Reference Downloads: 30
XML Downloads: 1519
Web Metrics
Days since publication: 3956
Overall contacts: 54930
Avg. contacts per week: 97.20
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2016): 12
Average cites per year: 1.20
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Scale dependency of the effects of landscape structure and stand age on species richness and aboveground biomass of tropical dry forests
vol. 16, pp. 234-242 (online: 23 August 2023)
Research Articles
The concept of green infrastructure and urban landscape planning: a challenge for urban forestry planning in Belgrade, Serbia
vol. 11, pp. 491-498 (online: 18 July 2018)
Research Articles
Evaluation of urban forest landscape health: a case study of the Nanguo Peach Garden, China
vol. 13, pp. 175-184 (online: 02 May 2020)
Research Articles
Communicating spatial planning decisions at the landscape and farm level with landscape visualization
vol. 7, pp. 434-442 (online: 19 May 2014)
Review Papers
Remote sensing-supported vegetation parameters for regional climate models: a brief review
vol. 3, pp. 98-101 (online: 15 July 2010)
Research Articles
How do urban dwellers react to potential landscape changes in recreation areas? A case study with particular focus on the introduction of dendromass in the Hamburg Metropolitan Region
vol. 7, pp. 423-433 (online: 19 May 2014)
Short Communications
The Polish landscape changing due to forest policy and forest management
vol. 2, pp. 140-142 (online: 30 July 2009)
Review Papers
Accuracy of determining specific parameters of the urban forest using remote sensing
vol. 12, pp. 498-510 (online: 02 December 2019)
Research Articles
Can the dynamics of forest restoration reduce landscape fragmentation in the Atlantic forest?
vol. 18, pp. 61-68 (online: 04 April 2025)
Review Papers
Remote sensing of selective logging in tropical forests: current state and future directions
vol. 13, pp. 286-300 (online: 10 July 2020)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







