Natural regeneration and species diversification after seed-tree method cutting in a maritime pine reforestation
iForest - Biogeosciences and Forestry, Volume 15, Issue 6, Pages 500-508 (2022)
doi: https://doi.org/10.3832/ifor4088-015
Published: Dec 14, 2022 - Copyright © 2022 SISEF
Research Articles
Abstract
Forest adaptation to global change has become one of the main objectives of forest management in recent years. Species and structural diversification by thinning is a well-known adaptation measure, often applied by managers in monospecific even-aged stands. However, regeneration fellings may be more efficient for enhancing naturalization and resilience of monospecific reforested pine stands, widely distributed throughout the Mediterranean basin. This work focuses on the evaluation of natural regeneration after seed-tree method cuttings, adopted as a diversification strategy in a 60-year old P. pinaster reforestation with varying presence of broad-leaved species (mainly Quercus spp.) in mountains of Southern Spain. A trial consisting of 78 circular plots (1 m radius) was installed to monitor annual tree-species regeneration over seven years, classified into seedlings (0-10 cm height) and saplings (10-130 cm). Scrub coverage and other ecological variables were also measured. Sampling was carried out in three stand types (Pure, Mixed 1 and Mixed 2, in order of increasing presence of broad-leaved species), according to forest species composition before the felling. Maritime pine regeneration was successfully achieved by the end of the study period (over 2000 trees ha-1 widely distributed throughout the stand), whereas Quercus spp. seedlings even colonised plots in which there were no seed trees. Pine seedling density was positively affected by summer precipitation and the presence of conspecific adult pines, and negatively influenced by scrub coverage. High temperatures during the mid-summer months (especially August) negatively affected the seedling to sapling change ratio as well as sapling survival during the study period. Our results point to regeneration fellings with retention of pine and broadleaf species as a suitable method for diversifying species composition in Pinus pinaster reforestations in the Mediterranean region.
Keywords
Adaptation, Global Change, Mixed Forests, Naturalization, Regeneration Fellings
Introduction
Monospecific conifer plantations have been commonly used for forest land restoration in many areas of the world ([36], [13]). In the Mediterranean region, the most frequent conifer species used in reforestations belong to the genus Pinus ([37]). Although the pioneer and fast-growing nature of most of the Mediterranean pine species facilitates the success of tree plantation, the structural homogeneity of these plantations caused by their monospecific and coetaneous character can increase their vulnerability to biotic and abiotic hazards ([23]). Moreover, some of these risks may be aggravated when reforestations cover large continuous areas ([37]).
Global change is threatening the stability of forest systems, with increasing risk of climate-driven disturbances and emergence of new pests and diseases ([46]), beyond the effect of other socioeconomic impacts, such as human migration or productivity changes ([5]). The Mediterranean basin is considered a climate change hotspot ([49]), where it is projected that ecosystems will suffer higher temperatures and a redistribution of rainfall, with less precipitation during winters leading to higher rainfall accumulation during spring and autumn ([19]). Consequently, an increase in forest fire risk ([54]), changes in plant phenology ([22]), alterations to both current and potential species distribution ([15]), or changes in shade tolerance, among others, have been identified as possible effects of the new climatic conditions. Measures to improve the adaptive capacity of Mediterranean pine reforestations to climate change are therefore urgent, as their vulnerability to both abiotic and biotic disturbances has already been shown ([45]).
Adaptation of forest management to climate change is already addressed in most of the policy agendas, such as the EU Forest Strategy, but it is still scarcely implemented in many forest types such as Mediterranean pine reforestations. Existing studies on the development of adaptive silviculture have pointed to the need for structural and specific diversification of both natural and reforested stands ([2], [53]). Silvicultural interventions, like disturbances influencing specific composition and forest structure, should be oriented towards these objectives ([4]). Thinnings have been broadly used to achieve structural ([32], [16]) and specific diversification ([47], [17]). Regeneration fellings can also be applied to enhance species diversification ([28]), which at some sites might be reinforced by artificial understory plantations ([34]).
The efficacy of forest management strategies for adapting forest systems to climate change has been recently revised in the context of the Mediterranean region ([52]). Among these strategies, thinnings were found to provide benefits in terms of short-term resistance, but the authors highlight the need for research into strategies that promote long-term adaptation, such as changes in tree-species composition. Therefore, further research is needed in the case of regeneration fellings, as a key phase in the conversion of monospecific even-aged plantations to stands with more complex structures ([35]). In this regard, the suitability of different regeneration systems for this conversion may depend on the specific ecological needs of existing species and on local environmental conditions, particularly climate conditions in the light of global change.
In this study we focus on the structural and species diversification of a Pinus pinaster reforestation in Southern Spain using the seed-tree method, a common regeneration system applied to this light-demanding species ([21], [42]). Maritime pine (Pinus pinaster Ait.) is a species widely spread throughout in the Mediterranean basin, thanks to the ready acclimation of the species and adaptation to a wide range of ecological conditions ([1]). In Spain, this species covers 600,000 ha of natural stands and more than 800,000 ha of reforested stands, most of these reforestations having been overtaken in the last 75 years due to an ambitious reforestation plan in the second half of the 20th century. This species is threatened by global change, which is increasing its vulnerability to both biotic (fungus infections, insects pests or nematodes) and abiotic damage (forest fires, droughts). In fact, a combination of both factors is already causing higher mortality rates in adult maritime pine stands in Spain ([18]). Hence, the conversion of maritime pine stands to more complex structures that would improve their resilience is now crucial.
Our main aims are to explore how the seed-tree method allows species diversification to be achieved depending on previous stand structure and species composition while ensuring regeneration of the main species (here P. pinaster), and how species regeneration is driven by weather conditions. For this purpose, we have studied natural regeneration over a seven year period in a P. pinaster reforestation located in a mountain area of Southern Spain (Sierra Morena). This pine reforestation was already partially diversified with broad-leaved species and was cut using the seed-tree method. Our hypotheses were: (i) the seed-tree method with retention of broad-leaved trees is valid to achieve sufficient pine regeneration (both in quantity and spatial distribution); (ii) this silvicultural method is also a suitable strategy for enhancing species diversification in maritime pine reforestations, despite scarce densities of broad-leaved species; and (iii) pine regeneration is promoted by mild-summer conditions and presence of conspecific trees.
Material and methods
Study area
The study was carried out in the “Arroyo del Azor” forest (38° 27′ 6″ N, 04° 20′ 14″ W) in Fuencaliente (Ciudad Real), in the Sierra Morena mountain range. We focused on a P. pinaster stand that was reforested by sowing between 1946 and 1949. Maritime pine is a native species in these mountains, as evidenced by the existence of small relic maritime pine stands nearby ([9]). Before the regeneration cuttings, the stand was dominated by 65-year old pine trees, with a variable density of broad-leaved trees in the understory (Quercus suber L., Quercus faginea Lam., Quercus pyrenaica Willd. and Arbutus unedo L.).
The study area covers 42.8 ha, most of which faces E-SE-S, with an average slope of 15%, ranging between 2% and 31%. Altitude ranges from 780 to 870 m a.s.l., with an average of 825 m. According to the FAO soil classification system ([12]), the soil is a haplic luvisol. The study site is characterized by average annual precipitation of 740 mm concentrated mainly in autumn and winter, and an average temperature of 15.3 °C, with a very hot, dry summer.
Experimental design and sampling methodology
The stem density of natural broad-leaved species was the key factor for the classification into forest stand types. First of all, we classified the area into “Pure” or “Mixed”, with Pure pine forest presenting above 90% in density of maritime pine, and Mixed forest a proportion of both pine and broad-leaved species above 10% in density ([11]). To distinguish between Mixed 1 and Mixed 2, according to a broadleaved tree presence criterion, we established a threshold at 70% of pine density ([20]), with “Mixed 1” between 70% and 90% of pine density, and “Mixed 2” below 70% (or more than 30% of broad-leaved trees). The three stand types were spatially mixed, as a result of an irregular spatial distribution of broad-leaved trees in the reforestation, due to natural colonization before and during the development of the pine trees.
Seed-tree method cuttings, applied in spring 2012, changed the forest structure, reducing pine densities significantly while retaining adult trees of broad-leaved species (Tab. 1). As seen in Tab. 1, species composition after the felling remained pure in “Pure” pine plots and mixed in “Mixed 1”, changing to broad-leaved pure in “Mixed 2” plots (pine density equal to 5%). The method applied consisted of using a harvester to cut and log pine trees, and a forwarder to pull them to the landing area. Crowns were also harvested and converted into chips for energy applications using a wood splitter.
Tab. 1 - Dasometric characterisation of study area per forest composition type, before and after the regeneration fellings. (Pt): P. pinaster; (Qs): Quercus suber; (Qf): Quercus faginea; Qp): Quercus pyrenaica; (Au): Arbutus unedo; (Brd): broad-leaved trees; (N): density, in trees per ha; (G): basal area, in m2 per ha; (d): mean diameter, in centimetres; (h): mean height, in metres; (Dc): crown diameter, in metres; (CR): crown ratio, in percentage; (*): Pt heights are dominant heights.
| Stand Type | Param | Pre-cuttings | Post-cuttings | Extracted (%) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pt | Qs | Qf | Qp | Au | Total | Pt | Qs | Qf | Qp | Au | Total | Pt | Brd | Total | ||
| Pure | N | 371.1 | 4.4 | 0 | 0 | 2.2 | 377.7 | 48.9 | 4.4 | 0 | 0 | 2.2 | 55.5 | 86.8 | 0 | 85.3 |
| G | 50.4 | 0.14 | 0 | 0 | 0.05 | 50.59 | 7.9 | 0.14 | 0 | 0 | 0.05 | 8.09 | 84.3 | 0 | 84 | |
| d | 41.1 | 20.2 | - | - | 16.3 | - | - | 20.2 | - | - | 16.3 | - | - | - | - | |
| h | 23.2* | 8.4 | - | - | - | - | - | 8.4 | - | - | - | - | - | - | - | |
| Dc | 6.5 | 4.4 | - | - | - | - | - | - | - | - | - | - | - | - | - | |
| CR | 32.1 | 65.9 | - | - | - | - | - | - | - | - | - | - | - | - | - | |
| Mixed 1 | N | 266.7 | 35.6 | 13.3 | 0 | 8.9 | 324.5 | 22.2 | 33.3 | 13.3 | 0 | 8.9 | 77.7 | 91.7 | 4 | 76.1 |
| G | 41.5 | 1.36 | 0.29 | 0 | 0.09 | 43.24 | 4.4 | 1.31 | 0.29 | 0 | 0.09 | 6.09 | 89.4 | 5.6 | 85.9 | |
| d | 45 | 20 | 15.2 | - | 11.3 | - | - | 20.2 | 15.2 | - | 11.3 | - | - | - | - | |
| h | 22.1* | 8.3 | 8.7 | - | - | - | - | 8.4 | 8.7 | - | - | - | - | - | - | |
| Dc | 7 | 4.5 | 5.6 | - | - | - | - | - | - | - | - | - | - | - | - | |
| CR | 36.8 | 65.9 | 73.6 | - | - | - | - | - | - | - | - | - | - | - | - | |
| Mixed 2 | N | 220 | 80 | 42.5 | 45 | 15 | 402.5 | 10 | 75 | 35 | 32.5 | 10 | 162.5 | 95.5 | 16.4 | 59.6 |
| G | 38 | 2.99 | 0.55 | 0.72 | 0.15 | 42.41 | 1.7 | 2.8 | 0.46 | 0.58 | 0.09 | 5.63 | 95.5 | 15.2 | 86.9 | |
| d | 46.8 | 21.2 | 12.7 | 14.1 | 11.5 | - | - | 20.9 | 12.5 | 14 | 10.7 | - | - | - | - | |
| h | 22.5* | 8.7 | 8 | 10.8 | - | - | - | 8.6 | 7.9 | 10.7 | - | - | - | - | - | |
| Dc | 7.7 | 5.5 | 5.1 | 4.2 | - | - | - | - | - | - | - | - | - | - | - | |
| CR | 37.2 | 67 | 70.3 | 76.7 | - | - | - | - | - | - | - | - | - | - | - | |
Broad-leaved trees were clearly below the pine canopy, as inferred from the differences in height between species (Tab. 1). Nevertheless, the crown development can be considered sufficient to produce fertile seeds for regeneration.
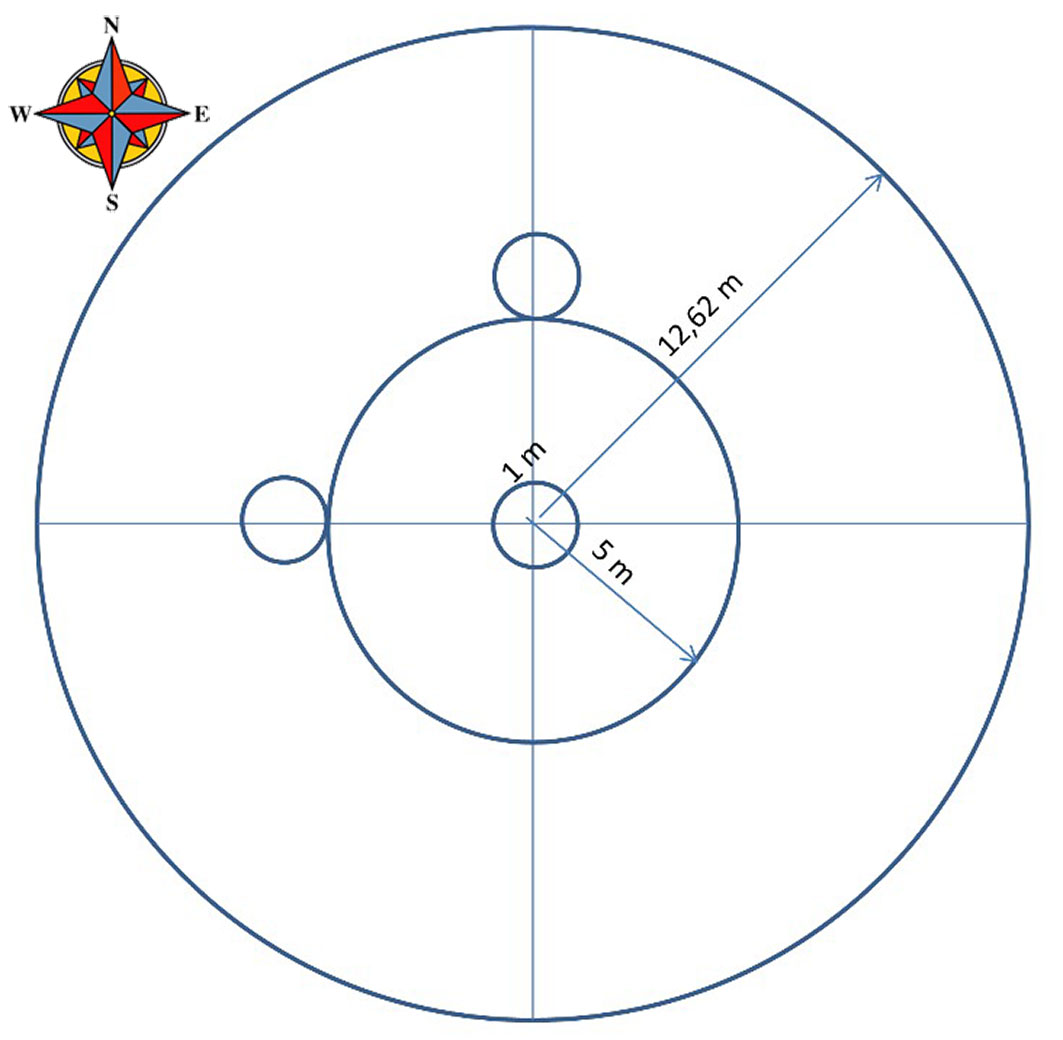
In 2011, prior to the cuttings, we established 26 circular inventory plots distributed among the three types of composition (9 plots in Pure, 9 in Mixed 1 and 8 in Mixed 2), arranged randomly in each group. In each circular plot of 12.62 m radius (500 m2), all adult trees of every species (>7.5 cm DBH, diameter at breast height) within the plot were located and their diameters measured. Height and crown dimensions were measured for the 5 thickest pine trees; while in the case of broadleaved species we selected the 2-4 thickest trees of each species to perform these measurements. Immediately after the cuttings, we recorded the standing mature trees to characterise the intensity of the seed-tree method cutting and the remaining stand. We completed the characterisation of each plot with the mensuration of some physiographic variables: slope, aspect and altitude.
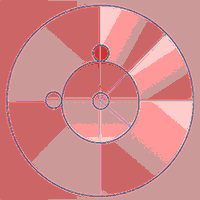
Regeneration (all plants smaller than 1.3 m in height) was recorded in three 1 m-radius plots installed within the inventory plots. These regeneration plots were systematically established, one being concentric to the inventory plot, and the other two at a distance of 6 m, north and west of the central plot (Fig. 1). Regeneration was measured every autumn from 2012, just after the fellings, until 2018. In these plots we also collected data for scrub (mainly Cistaceae species), litter and herbaceous coverage percentage in 2013, 2015, 2017 and 2018. Three regeneration plots from the Mixed 1 group were removed from the 2018 data due to the installation of bee hives which prevented access to these plots.
Fig. 1 - Size, shape and spatial distribution of inventory and regeneration plots.
To record the regeneration, we classified P. pinaster plants into seedlings (all plants smaller than or equal to 10 cm in height) and saplings (plants with a height > 10 cm). This classification filter was based on field observation, because all one-year plants (with no adult needles and lack of lignification) met the ≤10 cm height criterion and no >1-year-old plants were less than 10 cm in height. Quercus plants were not classified either by species or into seedlings-saplings, firstly because there were very few plants higher than 10 cm, secondly because hybridization hampered early, accurate, specific discrimination and finally, because the total number of plants was too small to apply significant statistical analysis differentiating Quercus species.
Climate data used in this study was provided by AEMET (Agencia Estatal de Meteorología), which has a meteorological station (5341C) located in Fuencaliente municipality, close to the study area. We selected maximum, minimum and mean monthly temperatures, in addition to monthly rainfall recordings for the entire study period (2012-2018). Using this data, we calculated seasonal mean, maximum and minimum temperatures, as well as seasonal precipitations.
Spatio-temporal descriptive analysis
We applied several descriptive statistical analyses to the regeneration data (seedlings and saplings of P. pinaster, and total plants of Quercus spp.) to explore the spatio-temporal dynamics of regeneration from the post-cutting situation to 2018.
Kendall’s correlation tests were applied due to the lack of normality in the data, and were carried out to show the dependence between plot pine densities of the two regeneration development stages in consecutive years (the six intervals between 2012 and 2018). We compared seedling density in year t with variation in sapling density between years t and t+1, leaving out plots where densities were zero in both years. This comparison requires the assumption of “no 2-year seedlings”, which means that all seedlings in one year either move to the sapling category or die.
To evaluate the success of the regeneration process for the three types of stand species composition, we analysed the percentage of plots where P. pinaster, Quercus spp., both species or no regeneration at all were present at the end of the studied period (2018). A χ2 analysis was performed to identify significant differences.
Furthermore, to determine whether there was any correlation between regeneration and development stages, we performed a Spearman’s correlation test on the 2018 regeneration records. For the purposes of this analysis, in accordance with the methodology described by Vergarechea et al. ([50]), we selected only those regeneration plots where at least one of the species was present, leaving out (0.0) combinations that could bias the results. In addition, to test the effect of adult tree species composition on presence/absence of P. pinaster/Quercus spp. regeneration, we applied contingency analysis using χ2 tables.
Finally, Spearman’s correlation tests were also applied to evaluate whether a relationship existed between the different development stage densities and any of the climatic variables (mean, maximum and minimum monthly average temperatures, and monthly precipitations) during the study period.
Modelling approach
To analyse the factors influencing the abundance of natural regeneration, we selected the regeneration density per plot as response variable of a generalized linear mixed model (GLMM). These models have been broadly used in regeneration studies ([26], [29]). As regeneration density is a variable which involves counting, a Poisson-distributed mixed model (with a log link function) provides a well-developed solution ([29]). The generic expression of the model can be expressed as follows ([43] - eqn. 1, eqn. 2, eqn. 3):
where yijk is the response variable (regeneration density per subplot j in a plot i and during a year k), λijk is the expectation of the Poisson distribution, x is a vector of explanatory variables, β is a column vector of maximum likelihood estimated coefficients and ui, uij and uk are the random effects.
Spearman’s correlation tests were applied as an exploratory analysis to filter the significance of all measured variables in seedling density. The variables that seemed to be correlated with seedling density, and that had been tested during the model development were those presented in Tab. 2.
Tab. 2 - List of variables used as independent variables in the fitting of the GLMM models, with a description and units, years of mensuration, mean and standard deviation (SD).
| Code | Description/Units | Years | Mean ± SD |
|---|---|---|---|
| Paug | Rainfall in August (mm) | 2013-2018 | 4.2 ± 3.8 |
| Psept | Rainfall in September (mm) | 2013-2018 | 33.5 ± 25.1 |
| PtotalSUM | Rainfall during summer (mm) | 2013-2018 | 38.8 ± 25.0 |
| Slope | Slope (%) | - | 15.5 ± 7.2 |
| Aspect | Factor reclassifying aspect in degrees into a [-1.1] scale (-1 = total shade; 1 = total sunlight) |
- | -0.56 ± 0.47 |
| Elevation | Elevation (m) | - | 822.2 ± 25.3 |
| Pre_Pt | P. pinaster pre-cuttings density (in trees plot-1) | 2011 | 14.4 ± 4.3 |
| Pre_GPt | P. pinaster pre-cuttings basal area (in m2 plot-1) | 2011 | 43.5 ± 9.2 |
| Post_Pt | P. pinaster density (in trees plot-1) | 2013 | 1.4 ± 1.4 |
| Post_Q | Quercus spp. density (in trees plot-1) | 2013 | 3.1 ± 3.3 |
| Post_GPt | P. pinaster basal area (in m2 plot-1) | 2013 | 4.7 ± 4.5 |
| Post_GQ | Quercus spp. basal area (in m2 plot-1) | 2013 | 1.8 ± 1.8 |
| Post_Ntotal | Total density (trees plot-1) | 2013 | 4.9 ± 2.9 |
| Post_Gtotal | Total basal area (m2 plot-1) | 2013 | 6.6 ± 4.1 |
| Scrub | Scrub coverage, standardized | 2013-2018 | 0.28 ± 0.27 |
| For_cod | Forest species composition (levels) | - | Pure, Mixed 1, Mixed 2 |
To avoid problems of independence with observations from the same plots, and to relate observations from the three regeneration plots to the inventory plot, we considered as a random effect in the model the regeneration subplot nested to the inventory plot. Year was also added as a random effect. In addition, to avoid problems with the different scales used for different variables, we standardized climatic and percentage variables using the square root transformation. Data from 2012 was discarded because of the almost null regeneration, which could puzzle our results.
In the model development, all variables were entered one by one and only those that were significant were selected. We then included the significant covariates in successive steps until there were no significant improvements in Akaike’s information criterion (AIC). After that, we added random effects to test whether they improved the accuracy of the model, and we also tested whether they met the normality assumption of random effects by performing Shapiro-Wilk tests. Finally, we looked for over or underdispersion problems in the selected model, and tested for this by adding an autoregressive factor.
We carried out the statistical analysis using the R software environment ([41]), and specifically packages “lme4” ([3]), “lmtest” ([55]) and “DHARMa” ([24]). The significance level was established at α=0.05.
Results
Temporal dynamics and climatic influences on the regeneration process
Regeneration of P. pinaster was more abundant than that of Quercus spp. in the three types of forest-species composition analysed during the 2012-2018 period (Fig. 2a, Fig. 2b). In 2012, immediately after the cuttings, average density for pines was very low, with few seedlings and only a small quantity of surviving pre-felling saplings.
Fig. 2 - (a) Average P. pinaster seedling (in continuous line) and sapling (in dashed line) densities per forest composition type and year. (b) Average Quercus spp. regeneration density per forest composition type and year.
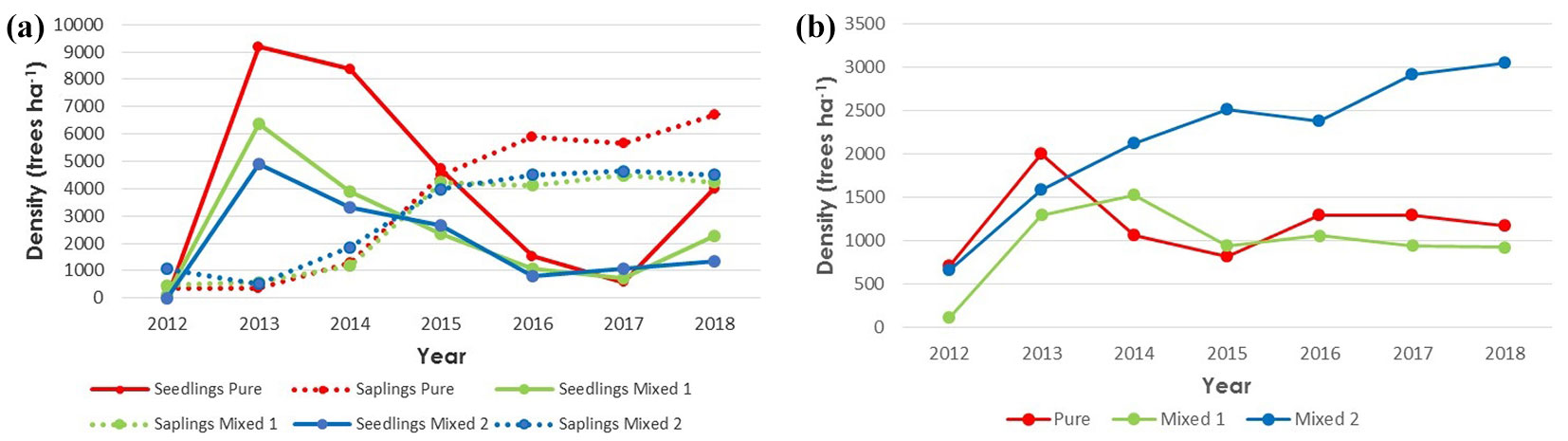
Since 2013, pine regeneration densities have been increasing, so that in 2018 there were average densities of 10,700, 6,500 and 5,800 trees ha-1 of total pine regeneration (in Pure, Mixed 1 and Mixed 2, respectively - Fig. 2a), with sapling percentages of 63%, 65% and 77%. Kendall’s correlation tests applied to seedling densities in year t and variations in sapling densities in year t+1 indicate that the establishment process (seedling to sapling transition) occurred between 2013 and 2016, as seedling densities per plot in years 2013 to 2015 showed strong positive correlation with variation in sapling density in years 2014 to 2016 (0.41, 0.46 and 0.44, respectively; p<0.05).
Quercus spp. dynamics were slightly different. In 2012, the regeneration density of oak was almost null in Mixed 1 type, while Pure and Mixed 2 types presented less than 1000 trees ha-1 of oak regeneration, a part of which were probably pre-felling survivors. A growth in oak densities in all forest types was observed in 2013, but the dynamics changed after that. Pure plots reached a maximum density, followed by a large fall in 2014, after which the situation has stabilized in those plots. Oak regeneration increased in Mixed 1 plots in 2014 but density fell in 2015 and then stabilized until 2018. Densities in Mixed 2 plots, however, increased gradually, reaching a maximum in 2018 (Fig. 2b).
Despite the relatively abundant presence of adult trees of Arbutus unedo, no regeneration plants of this species were found in any of the plots during the study period.
Spearman’s correlation test, applied to identify whether any significant relationship could be found between regeneration densities (by species and development stages) and climatic variables, showed that total summer precipitation (Spearman’s r = 0.81, p = 0.048) and especially September precipitation (r = 0.86, p = 0.03) was strongly associated with seedling survival in summer. These tests revealed significant negative relationships between August (r = -0.9, p = 0.015) and September (r = -0.82, p = 0.048) average maximum temperatures and changes in sapling densities, indicating that summers with higher temperatures present fewer seedling-to-sapling transitions and higher seedling and sapling mortality.
Descriptive spatial analysis
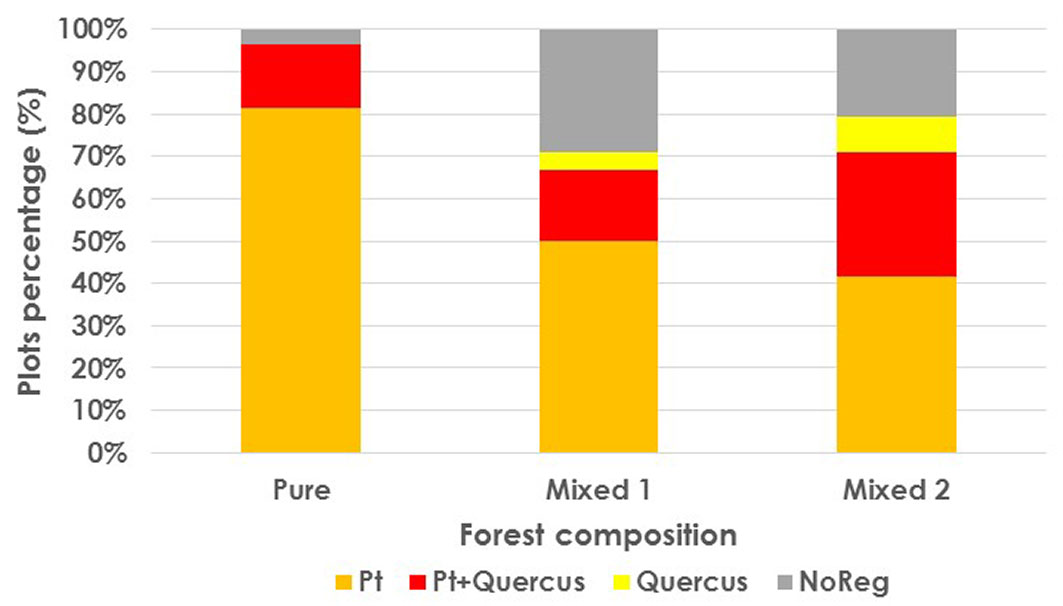
P. pinaster regeneration was well distributed throughout the stand at the end of the monitoring period (2018). In Pure pine forest, almost all the plots (96.3%) had pine regeneration, while Quercus was only present in 14.8% of the plots (Fig. 3). As regards regeneration development stages, 81.4% of plots had saplings (37% of them having only saplings and the remaining 44.4% both seedlings and saplings). The percentages in Mixed 1 plots were quite different. A large percentage of plots (29.2%) had no regeneration at all. Pine regeneration was present in 66.7% of plots (all of them with sapling presence), while Quercus plants were only present in a scant 20.9% of plots. (Fig. 3). Finally, in the Mixed 2 forest structure, pine presence was similar to that of the Mixed 1 structure (70.9%, 58.3% saplings), although fewer plots had no regeneration at all (20%) and Quercus regeneration was present in 37.5% of plots (Fig. 3). A χ2 test was performed for the percentage distribution in all the forest species composition types, indicating that there were significant differences among the percentage vectors in different types of forest composition.
Fig. 3 - Occurrence frequencies of P. pinaster and Quercus spp. regeneration in 2018 by forest composition. (Pt): P. pinaster; (Pt+Quercus): plots with both species; (NoReg): plots where no regeneration was found. A χ2 test showed that frequency distribution was significantly different between all types of forest composition.
A Spearman’s rank correlation coefficient test to determine whether a relationship existed between Pinus and Quercus spp. and development stages at plot level in 2018, pointed to significant negative correlation between P. pinaster seedlings and Quercus spp. regeneration, but no correlation between P. pinaster development stages or to P. pinaster saplings and oak regeneration were found (Tab. 3).
Tab. 3 - Spearman’s correlation matrix between plant densities in each development stage during the last year of the study (2018). (*): p<0.05. Correlation tests were run with 43 plots for seedlings-Quercus, 62 plots for seedlings-saplings and 60 plots for saplings-Quercus.
| Species (stage) | P. pinaster (seedlings) |
P. pinaster (saplings) |
Quercus spp. |
|---|---|---|---|
| P. pinaster (seedlings) | 1 | - | - |
| P. pinaster (saplings) | -0.03 | 1 | - |
| Quercus spp. | -0.39* | -0.11 | 1 |
Tab. 4 shows the association between the presence/absence of natural regeneration of P. pinaster (both seedlings and saplings) and Quercus, and the composition of the adult tree stratum. Presence of maritime pine seedlings is significantly higher in Pure stands. Forest composition type does not have a significant influence on the presence/absence of pine saplings or Quercus regeneration.
Tab. 4 - Contingency analysis for the composition of the adult tree strata and presence/absence of regeneration in 2018. Numbers in italics are the expected values.
| Species | Stage | Presence/ Absence |
Pure | Mixed 1 | Mixed 2 | Total | χ2[2] | p-value |
|---|---|---|---|---|---|---|---|---|
| P. pinaster | Seedlings | Presence | 16 | 7 | 7 | 30 | 6.5201 | 0.0383 |
| 10.8 | 9.6 | 9.6 | ||||||
| Absence | 11 | 17 | 17 | 45 | ||||
| 16.2 | 14.4 | 14.4 | ||||||
| Saplings | Presence | 22 | 15 | 15 | 52 | 2.9282 | 0.2313 | |
| 18.7 | 16.6 | 16.6 | ||||||
| Absence | 5 | 9 | 9 | 23 | ||||
| 8.2 | 7.3 | 7.3 | ||||||
| Quercus spp. |
Total | Presence | 4 | 5 | 9 | 18 | 3.7788 | 0.1512 |
| 6.5 | 5.8 | 5.8 | ||||||
| Absence | 23 | 19 | 15 | 57 | ||||
| 20.5 | 18.2 | 18.2 |
Modelling regeneration
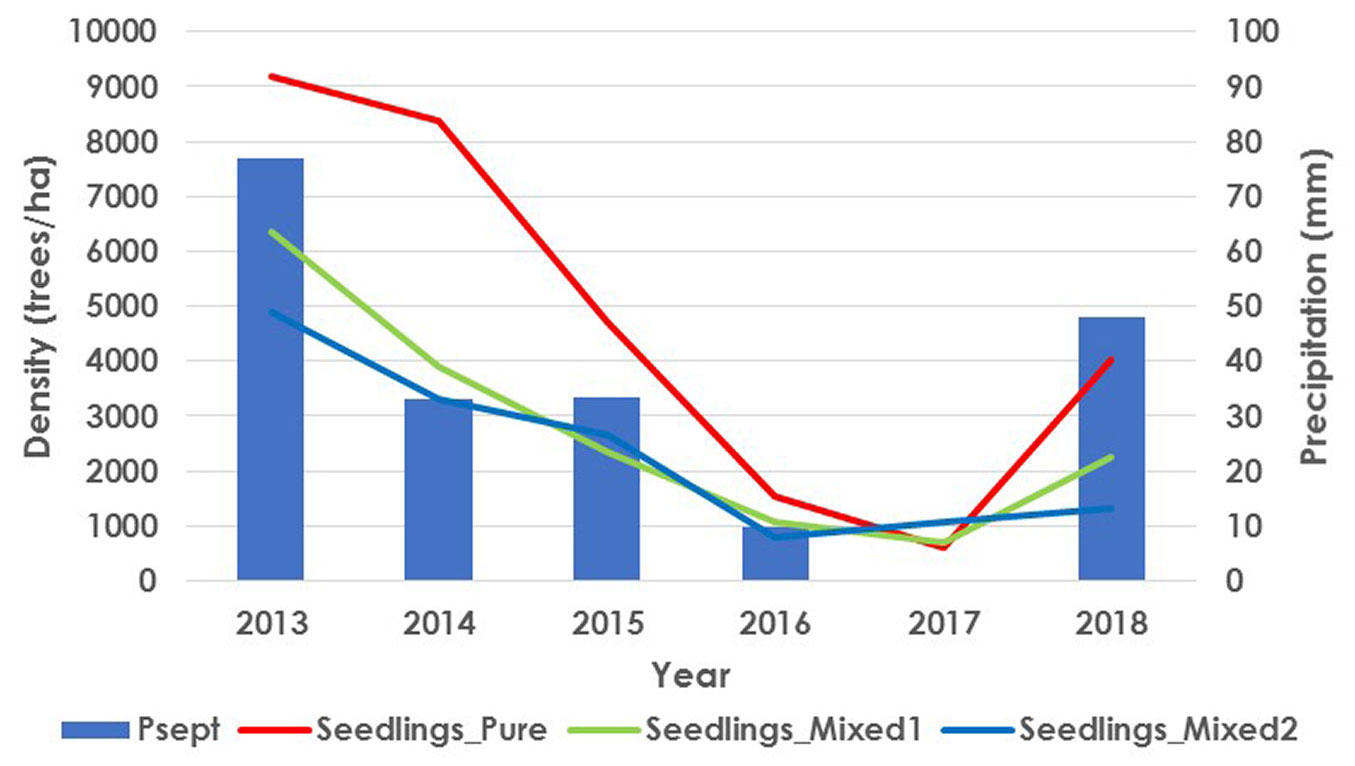
The observed records confirmed the accuracy of the fitted model for pine seedlings. Subplot/Plot, Plot and Year random effects improved the model and had a variance of 0.45 (Subplot/Plot effect), 0.07 (Plot effect) and 0.05 (Year effect). The model explained 59.1% of the total variance of the seedling density. Entering an autoregressive factor in the model had no significant influence on the accuracy of the model. The selected model was (eqn. 4):
with Psept being the September precipitation (mm), Scrub is the scrub coverage, Mixed 1 and Mixed 2 are the forest composition types, and u as the random effects (ui=Plot, uij=Subplot/Plot, and uk=Year). Tab. 5 shows the maximum likelihood coefficients (β), their significance and standard errors.
Tab. 5 - Coefficients estimated by maximum likelihood used in the model for pine seedlings, with standard error and p-value. (Psept): rainfall in September (mm), squared-root transformed; (Mixed 1, Mixed 2): types of mixture; (Scrub): scrub coverage, standardized.
| Effect | Parameter | Estimate | Std. error | p-value |
|---|---|---|---|---|
| Intercept | β 0 | -1.0790 | 0.3207 | 0.0008 |
| Psept | β 1 | 0.2390 | 0.0420 | <0.0001 |
| Mixed 1 | β 2 | -0.5846 | 0.2512 | 0.0200 |
| Mixed 2 | β 3 | -0.7740 | 0.2624 | 0.0032 |
| Scrub | β 4 | -1.0012 | 0.3634 | 0.0059 |
We observed a positive effect of September precipitations on pine seedling densities, which can also be observed in Fig. 4. Scrub was found to have a significant negative effect, while forest composition was observed to affect seeding density, with higher seedling density in Pure pine plots than in Mixed plots, this effect also being apparent in the contingency analysis presented in Tab. 4.
Fig. 4 - Seedling density per year compared with September rainfall.
Tab. 6 - Coefficients estimated by maximum likelihood used in the Quercus spp. regeneration model, with standard error and p-value. (Post_GQ): basal area of Quercus spp. after the cuttings.
| Effect | Parameter | Estimate | Std. error | p-value |
|---|---|---|---|---|
| Intercept | β 0 | -2.2568 | 0.3665 | <0.0001 |
| Post_GQ | β 1 | 0.2761 | 0.1212 | 0.0227 |
Quercus spp. regeneration was strongly related with Quercus basal area of adult trees after the fellings (Post_GQ). In this case, the Subplot/Plot random effect had significant influence in the model performance, but not the Year. The model explained 50.4% of the total variance of Quercus density. Tab. 6 shows the maximum likelihood coefficients (β), their significance and standard errors for this model (eqn. 5):
No accurate models were found for pine saplings.
Discussion
In this study we identified key factors driving the regeneration process of pine and broadleaf species in a maritime pine reforestation. This information is crucial to design silvicultural treatments for converting even-aged pine monocultures to complex forests. Forest management aimed at promoting natural regeneration of planted forests must be considered in order to increase plant biodiversity in the face of climate change, given that current research points to greater benefits of more diverse forests in comparison to monocultures ([31]). This is of critical importance for species which have been planted over large areas, as in the case of Pinus pinaster in South-Western Europe and Northern Africa ([1]). The urgency is even greater in Spain where most of these plantations are reaching rotation age.
Our results point to the successful regeneration of maritime pine throughout the study area, which is in accordance with the pioneer character of this species. Average pine densities were greater than 2000 trees ha-1 in all forest composition types in 2018, this figure being the minimum density suggested to indicate successful pine regeneration at the end of the regeneration period ([39]). In fact, sapling densities above 4000 trees ha-1 were found in all forest composition types (with a maximum of 6700 trees ha-1 in Pure pine plots), indicating adequate establishment of pine seedlings to ensure the success of the pine regeneration process. Moreover, the sapling presence percentages in the plots, ranging from 58% in Mixed 2 plots to 81.7% in Pure plots, reflect a good spatial distribution of established pine regeneration.
As regards the broad-leaved species regeneration, the observed negative correlation between the density of pine and Quercus regeneration (Tab. 3) could be due to differences in establishment requirements of the species, as suggested by some authors ([50]), with maritime pine being a shade intolerant species while Mediterranean oaks show higher shade tolerance. In the case of cork oak, our results support those of previous studies which point to the need for adult-tree cover in the first stages of regeneration and subsequent progressive release of this cover to enhance the growth of established plants ([48], [8]). Pausas et al. ([38]) found that oak regeneration was positively related to pine basal area in a cork oak-maritime pine mixed forest. However, our findings showed a positive relationship between oak regeneration and basal area of adult broad-leaved trees, but not with basal area of adult pines, suggesting the importance of both factors, i.e., protective canopy cover and presence of seed trees for oak regeneration.
In the study area, regeneration fellings involved intense canopy cover removal (Tab. 1), contrary to the reported requirements of Quercus species, which may explain the low presence of oak. However, the different dispersal methods of the species (anemochory in pine, zoochory and gravity in oaks) could also explain the differences in observed distribution of the species. Mixed 2 plots, with more Quercus spp. seed trees, presented a higher oak regeneration, both in terms of density (more than 3000 trees ha-1) and spatial distribution (37.5% of plots with oak regeneration). However, it is worthy of note that plots without significant presence of adult Quercus trees now present high densities of oak seedlings, which indicates that species diversification is taking place. In this regard, the proximity of the different forest structures, which leads to ecotone-like situations, may have influenced our results.
The analysis of the effect of ecological variables (topographic, climatic or stand) on the regeneration of both species supported the hypothesis that summer conditions are fundamental in the development of the regeneration process. On one hand, we found that September precipitation has a significant positive influence on summer survival of seedlings. As P. pinaster seeds mainly germinate in spring ([44]), late summer rainfall is crucial for seedling survival and recovery from intense summer drought, and may also induce autumn seedling incorporation. Our results agree with those of Ruano et al. ([42]) and Rodríguez-García et al. ([40]), who reported positive effects of summer precipitation on seedling germination and initial development in natural P. pinaster forests in the Spanish Northern Plateau. Calama et al. ([6]) reported that summer rainfall and progressive crown canopy release are the key factors assuring maritime pine regeneration, while Fernandes et al. ([14]) found that seedlings emerging in spring were more likely to die than those emerging in autumn, perhaps due to summer drought and to the negative influence of a warm-summer climate on survival rate in Portugal. Furthermore, we found a negative effect of maximum average temperatures in August and September on seedling-to-sapling transition and on sapling survival. This effect was also recorded in studies by Moreno-Fernández et al. ([33]) and Vergarechea et al. ([51]), both conducted in the Spanish Northern Plateau.
These findings regarding the significance of summer precipitation and temperature will be of increasing importance as drought periods become longer and more intense due to climate change, possibly leading to a bottleneck in the regeneration process. Furthermore, this increasing duration and intensity of summer drought due to lower levels of precipitation and more stressful temperatures could lead to plant stress and a rise in infections by pests or diseases, e.g., the potential increase in infections by bast scale (Matsucoccus feytaudi), the effects of which are currently negligible in the Iberian Peninsula but severe in Italy and South-Eastern France ([27], [30]), or an increase in the distribution area of the pine wood nematode (Bursaphelenchus xylophilus Steiner & Buhrer 1934) in the Iberian Peninsula ([10]).
Herbaceous and scrub coverage has also been identified as another factor affecting tree regeneration. Herbaceous cover was almost entirely absent and only appeared before the third year, so the negative effect of this factor reported in the literature ([21], [39]) was not found at our study site. However, scrub cover had a significant negative effect (competition) in the pine seedling model. This negative competition effect of scrub has also been highlighted in the literature, e.g., after forest fires ([7]). Maltoni et al. ([30]) reported that patches with high levels of maritime pine regeneration along with shrubs can provide protection against browsing for oak regeneration, this being a possible strategy in areas with high pressure from wildlife. In this regard, there is a large wildlife population in our study area (mainly deers, roe deers and wildboars) with frequent browsing damages observed in regeneration plants. Hence, the fact that a significant negative effect of scrub species presence was identified suggests that competition from the scrub layer may be more detrimental to pine regeneration than herbivore browsing.
Forest composition and structure of adult trees are also important factors influencing natural regeneration. We observed that presence of conspecific trees (pine or oaks) stimulated regeneration density of both species. These results are supported by those of several other studies, which indicate that P. pinaster requires a certain degree of canopy cover in Mediterranean environments to mitigate the impact of summer drought on plant survival and establishment ([33], [50]). González-Alday et al. ([21]) and Ruano et al. ([42]) suggested an interval of 25%-50% in basal area removal as the optimum for pine seedling survival on sandy soils in the Northern Plateau of Spain. These studies, which would appear to contradict the shade intolerant behaviour attributed to P. pinaster ([33]), suggest that this species may benefit from a degree of coverage in order to survive the high summer temperatures and droughts, which are frequent in Mediterranean environments. As summer droughts can be a bottleneck for forest regeneration, forest managers should consider retaining some of the adult pine trees to protect regeneration, bearing in mind that these trees could also act as seed and biodiversity reservoirs ([25]). Seed production and seed dispersal, factors not analysed in this study, may also contribute towards explaining differences in seedling densities.
Conclusions
The application of the seed-tree method with retention of broad-leaved species was found to be a valid regeneration approach to promote both pine regeneration and species diversification in reforested maritime pine stands, even though very few adult trees of broad-leaved species were present. Although P. pinaster is a shade-intolerant species that requires substantial canopy release, retaining some canopy cover may favour pine seedling survival. Hence, the seed-tree method would appear to be preferable to clear-cuttings under Mediterranean climates with severe hot and dry summers, conditions that are likely to become more frequent due to climate change. The presence of shade-tolerant species such as oaks may increase in mixed stands, thus diversifying forest composition as long as adult trees of these species are not extracted during the regeneration fellings until sufficient regeneration has been promoted. Strict forest management plans aimed at adaptation to global change, should be put into practice in vulnerable forest stands such as pine reforestations. It is expected that the findings of this research will aid forest managers in the application of adaptive silviculture regeneration techniques in reforestations, ensuring forest persistence and promoting species diversification.
Acknowledgements
The authors wish to thank Andrés Bravo Oviedo, Eduardo López Senespleda and Sven Mutke for their collaboration in the fieldwork, the Forest Service of Ciudad Real province for their contribution to the development of this study, and AEMET for providing climate data. This study has been financed by the following projects: AGL20 11-29701-C02-01, AGL2016-77863-R (FORADMIT), and AGL2017-83828-C2.1R. SDF has also received funding from a FPU predoctoral contract of the Spanish Ministry of Education (FPU18/04597).
References
Gscholar
CrossRef | Gscholar
Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
José Alfredo Bravo-Fernández 0000-0001-5909-3131
Sonia Roig-Gómez 0000-0002-2057-3020
Departamento de Sistemas y Recursos Naturales, Escuela Técnica Superior de Ingeniería de Montes, Forestal y del Medio Natural, Universidad Politécnica de Madrid, c/ José Antonio Novais 10, 28040 Madrid (Spain)
Ricardo Ruiz-Peinado 0000-0003-0126-1651
Instituto de Ciencias Forestales - ICIFOR, INIA-CSIC, crta. de La Coruña, km. 7.5, 28040 Madrid (Spain)
Corresponding author
Paper Info
Citation
De Frutos S, Bravo-Fernández JA, Roig-Gómez S, Del Río M, Ruiz-Peinado R (2022). Natural regeneration and species diversification after seed-tree method cutting in a maritime pine reforestation. iForest 15: 500-508. - doi: 10.3832/ifor4088-015
Academic Editor
Susanna Nocentini
Paper history
Received: Feb 27, 2022
Accepted: Oct 24, 2022
First online: Dec 14, 2022
Publication Date: Dec 31, 2022
Publication Time: 1.70 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2022
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 28694
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 22211
Abstract Page Views: 4051
PDF Downloads: 1946
Citation/Reference Downloads: 8
XML Downloads: 478
Web Metrics
Days since publication: 1288
Overall contacts: 28694
Avg. contacts per week: 155.95
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2022): 2
Average cites per year: 0.50
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Optimizing silviculture in mixed uneven-aged forests to increase the recruitment of browse-sensitive tree species without intervening in ungulate population
vol. 11, pp. 227-236 (online: 12 March 2018)
Research Articles
Approaches to classifying and restoring degraded tropical forests for the anticipated REDD+ climate change mitigation mechanism
vol. 4, pp. 1-6 (online: 27 January 2011)
Research Articles
Tree-oriented silviculture: a new approach for coppice stands
vol. 9, pp. 791-800 (online: 04 August 2016)
Research Articles
Seeing, believing, acting: climate change attitudes and adaptation of Hungarian forest managers
vol. 15, pp. 509-518 (online: 14 December 2022)
Review Papers
Problems and solutions to cork oak (Quercus suber L.) regeneration: a review
vol. 16, pp. 10-22 (online: 09 January 2023)
Review Papers
Should the silviculture of Aleppo pine (Pinus halepensis Mill.) stands in northern Africa be oriented towards wood or seed and cone production? Diagnosis and current potentiality
vol. 12, pp. 297-305 (online: 27 May 2019)
Review Papers
Opportunities for coppice management at the landscape level: the Italian experience
vol. 9, pp. 775-782 (online: 04 August 2016)
Research Articles
Modelling natural regeneration of Oak in Saxony, Germany: identifying factors influencing the occurrence and density of regeneration
vol. 16, pp. 47-52 (online: 16 February 2023)
Research Articles
Methods for predicting Sitka spruce natural regeneration presence and density in the UK
vol. 12, pp. 279-288 (online: 23 May 2019)
Research Articles
Long-term dynamics of stand structure and regeneration in high-stocked selection fir-beech forest stand: Croatian Dinarides case study
vol. 14, pp. 383-392 (online: 24 August 2021)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







