Suitability of Fagus orientalis Lipsky at marginal Fagus sylvatica L. forest sites in Southern Germany
iForest - Biogeosciences and Forestry, Volume 15, Issue 5, Pages 417-423 (2022)
doi: https://doi.org/10.3832/ifor4077-015
Published: Oct 19, 2022 - Copyright © 2022 SISEF
Research Articles
Abstract
European beech (Fagus sylvatica L.) is the most important tree species in Central Europe and is considered to be relatively resistant to climate warming. However, dry summers in the last five years led to considerable damage in beech stands in Southern Germany (SG). Assisted migration of drought resistant beech provenances including those of Oriental beech (Fagus orientalis Lipsky) may help to stabilise Central European beech forests under climate change. The focus of this study is to compare the climatic ranges of F. sylvatica and F. orientalis using quantile distribution of climatic variables based on WorldClim data at forest sites within their natural distribution area. Temperature, precipitation, and aridity quantile ranges showed that F. orientalis is better adapted to warmer and drier climate compared to F. sylvatica. The quantile distribution method was applied to the whole range of the species to map the habitat suitability for both species at marginal sites in the target region (SG) in the current climatic scenario (1970-2000) and in a warmer scenario (+2°C) using the climate marginality index (CMI), i.e., the distance of sites to the xeric edge at low-latitude and low-altitude distribution limits for the species. To this purpose we applied the simple BIOCLIM algorithm using annual temperature and precipitation as climatic variables. According to our results, F. orientalis seems a promising species with a high potential for future afforestation activities in Southern Germany, especially at marginal sites of European beech forests. However, before introducing F. orientalis on a larger scale in the study area, further research on the species ecology and genetics are needed. For further application of the quantile range method, we produced tables of the vigintiles of the climatic range for both species, which can be used for estimating CMI based on WorldClim data in other regions.
Keywords
BIOCLIM, Climate Change, Climatic Marginality Index, Climatic Range, Ellenberg Quotient, Macroecology, Quantile Distribution, Species Distribution
Introduction
European beech (Fagus sylvatica) is the major tree species in Central European forests ([15], [3]) and was previously considered to be comparatively resistant against climate warming in Germany ([47]). However, dry summers in recent years led to considerable damage in beech stands in Southern Germany (SG). Notably, beech is fundamental in the economically viable close-to-nature silviculture to be applied in stands with high conifer proportion in Central Europe ([15], [47], [5]). For maintaining a large share of beech forests, an assisted migration of drought resistant beech provenances or the closely related Fagus orientalis ([38]) are major options to be studied.
This pilot study lies within the framework of the project sensFORclim, and aimed to explore the suitability of F. orientalis in Southern Germany through macroecological data (descriptive statistics, climate envelopes) with a focus on F. sylvatica forest sites. In sensFORclim climate resilient populations of main tree species are analyzed by various methods (climate envelopes, dendroecology, physiology, and genetics) and at different spatial scales (continent, region, population). The target regions of our study are four federal states in Southern Germany (Baden-Wuerttemberg, Bavaria, Thuringia, Saxony).
Both the sister species F. sylvatica and F. orientalis are major tree species in their natural distribution area ([25], [28]). They are very shade-tolerant, dominant species preferring loamy soils and a fresh water regime, although they occur across a wide range of soils with widely differing texture class, soil acidity and nutrient supply ([15], [16], [46], [28], [25]). Yet, the distribution patterns of the species in the south-eastern Balkans suggest that F. orientalis may occur on drier and warmer sites than F. sylvatica ([28]).
Although much information on the ecology and genetics of both species is available ([11], [8], [23], [39], [2]), a comparison of the suitability of both species based on quantitative climate data is still missing, especially regarding the suitability of F. orientalis at marginal sites of F. sylvatica in Southern Germany.
In this study mapping techniques focusing on the rear edges of the species distribution ([19]) were used to estimate the climatic marginality index (CMI), which measures the distance of a site to the warm and dry niche edges of a species (i.e., the xeric limits - [32]). The simple mapping algorithm of the package BIOCLIM ([7], [4]) was used to model species distribution based on quantile range. CMI estimates represent the suitability of the species as compared to conventional niche models, whose predictions of low suitability cannot be associated to the rear or leading edge ([31], [32]).
The aim of this study was to assess the current and future suitability of F. sylvatica and F. orientalis in Southern Germany based on CMI estimated through the BIOCLIM algorithm using historic (1970-2000) climate data ([17]) and the predictions of a warmer climatic scenario (+2 °C) over the period 2071-2100.
Material and methods
Climate variables
To describe the species’ climatic ranges, we used 6 bioclimatic variables from the WorldClim data set ([17]), and two additional derived climatic variables. Bio1 (annual mean temperature, Tyr) and Bio12 (annual precipitation, Pyr) are fundamental climate variables which have already been used for modelling the distribution of beech species ([16], [32]). Bio5 (max temperature of warmest month, Tmax), Bio6 (min temperature of coldest month, Tmin), Bio10 (mean temperature of warmest quarter, Twq) and Bio18 (precipitation of warmest quarter, Pwq) are further important parameters for predicting the probability of occurrence of a species based on real observations ([31], [45]). As an additional parameter, we calculated the continentality index (Conti) after Conrad ([10]), which was successfully applied for estimating tree species distributions ([45] - eqn. 1):
where A is the difference between Bio5 and Bio6 and Φ is the geographic latitude. Following Andrade & Corte-Real ([1]), sites can be classified in 5 climatic categories based on Conti (in %): -20 / 20 = hyper-oceanic; 20 / 50 = oceanic/maritime; 50 / 60 = sub-continental; 60 / 80 = continental; 80 / 120 = extreme/hyper-continental climate. We also calculated the Ellenberg Quotient (EQ, [15]), a common aridity index often applied not only in Central Europe ([16], [43], [32], [24]), which is computed from July temperature (T07) and annual precipitation (Pyr - eqn. 2):
As the xeric limit is approached, the EQ values increase ([32]). EQ indicates the dominance of European beech in Central European forests ([15]). Beech forests with low proportions of other tree species prevail where EQ is in the range of 10 to 20. Lower EQ values (around 10) are typical for beech-fir forests. Subalpine beech forests usually occur at EQ < 10. At sites with EQ values between 20 and 30 beech forests prevail, while an increase in EQ is typical of forests with higher proportion of oak. Oak forests without beech are usually found at sites with EQ > 30 ([15]).
For assessing possible future range shifts of both the study species, we assumed a simple warming scenario (Tyr +2°C) with no changes in annual precipitation. This corresponds approximately to the RCP4.5 scenario for Southern Germany for the period 2071-2100 ([14]).
Species data
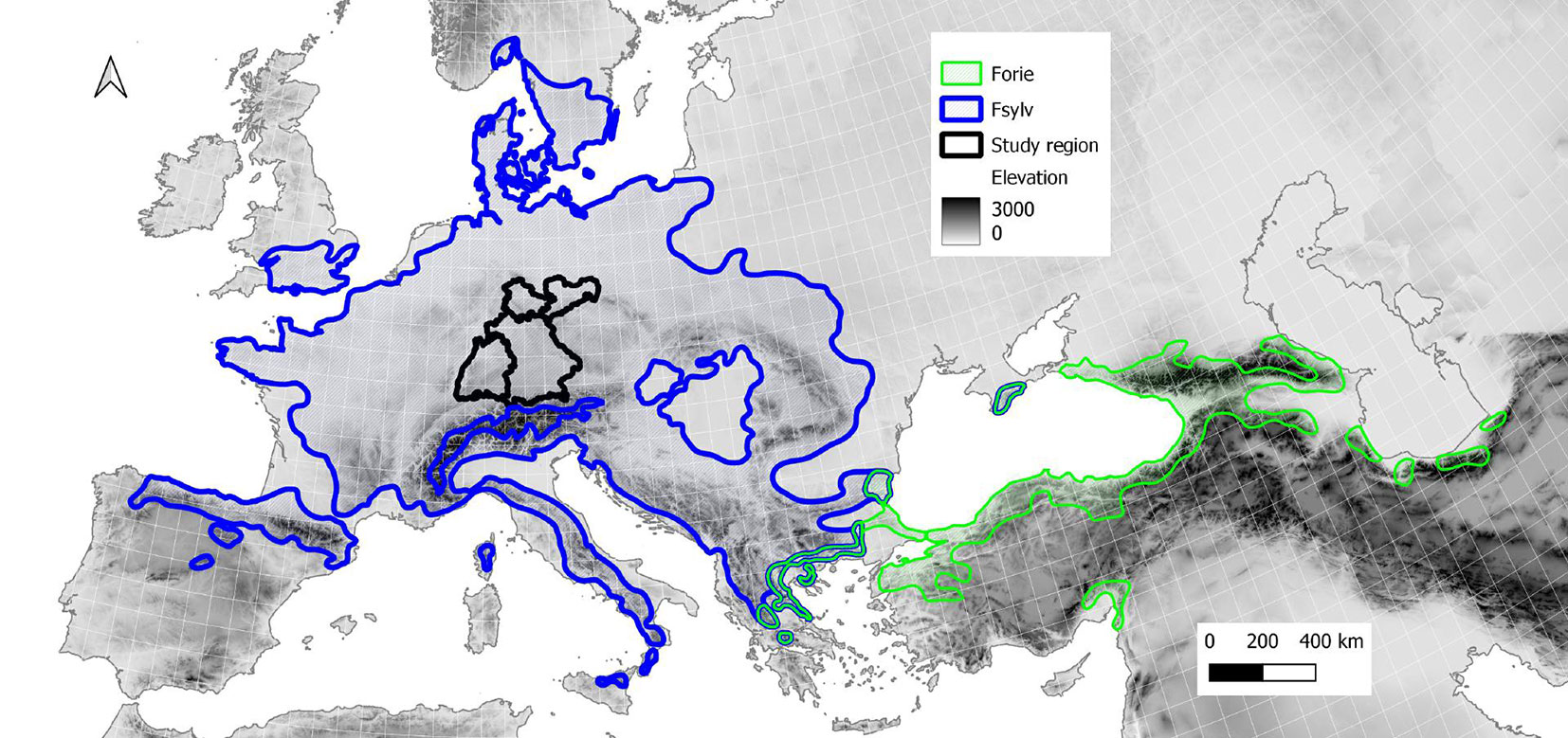
The entire distribution areas of both F. sylvatica and F. orientalis (Fig. 1) were derived from Caudullo et al. ([9]) and the forest layer map of COPERNICUS with approx. 100 m resolution (GLC100 v.3 2015), and used for modeling the potential beech range ([6]). The forest type layer provides discrete values for each type of forest and for all pixels where the tree (forest) cover fraction exceeds 1%. We selected pixels belonging to the deciduous broadleaf and mixed forests categories within the distribution area according to Caudullo et al. ([9]) based on a 4×4 km grid. Using this approach we gathered 37.618 records for F. sylvatica and 7.624 records for F. orientalis. The target area in Southern Germany included 2048 records. To obtain systematic and equidistant records, spatial data were processed using the projection ETRS89-extended / Europe Lambert-Azimuthal Equal Area (LAEA - ESPG = 3035).
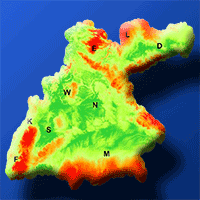
Fig. 1 - Distribution area of F. sylvatica (Fsylv - blue outline) and F. orientalis (Forie - green) according to Caudullo et al. ([9]), and the target area of this study in Southern Germany (black outline) consisting of four German states (Baden-Wurttemberg, Bavaria, Saxony, Thuringia). Background shading: elevation grid from WordClim ([17]).
Climatic ranges
The comparison of both species was carried out based on the quantile distribution of each climatic variable over the entire species range using boxplots, a basic descriptive statistical method. The interquartile range of each climatic variable (25%-75% of the univariate distribution) can be interpreted as the interval of preferable conditions along the distribution range of a species ([33]). Boxplots are a univariate equivalent to simple climate envelopes based on presence-only data in the conventional species distribution modelling ([7], [4]). Such techniques are often preferred in situations where real absence or presence data is missing ([18]).
Mapping
To map the suitability of both species in the target region (SG), we derived the quantile distribution of each climatic variable considered from the whole climate range of both species (Fig. 2). We obtained the quantiles of Tyr an Pyr using the BIOCLIM algorithm ([7]). BIOCLIM is closely related to Hutchinson’s idea of the species niche as a hypervolume ([27], [4]) and is perhaps the simplest approach to estimate the climatic envelope ([18]) as it is based on descriptive statistics. Despite its simplicity, it can keep up with inferential models when compared with mechanistic models ([21]). The CMI can be derived from BIOCLIM outputs by sorting the quantiles from the rear edge (the xeric limit, with high temperature and low rainfall) versus the leading edge (with low temperature and high precipitation - Hampe & Petit ([19]). The quantile distribution of the entire climate range of both species was applied to current and future climate data in the target region using the empirical cumulative distribution function (“ecdf”) in R ([41]). The model output represents the minimum CMI of both variables according to Liebig’s principle ([22]).
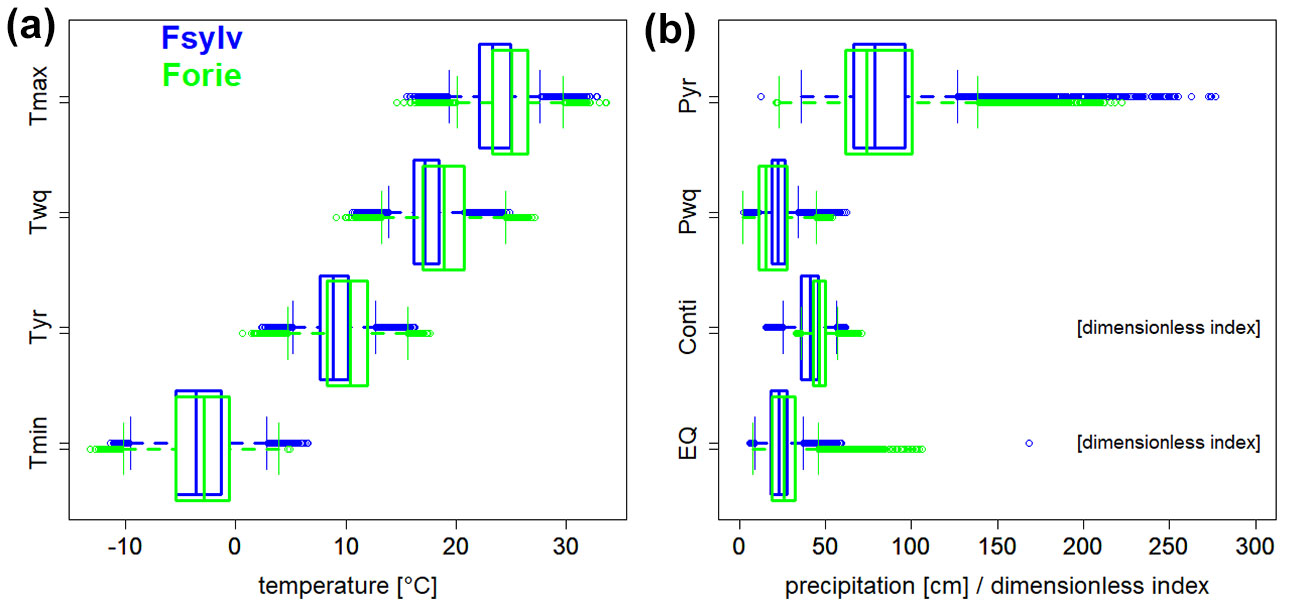
Fig. 2 - Climatic ranges of Fagus sylvatica (Fsylv) and F. orientalis (Forie) according to four temperature based bioclimatic variables (a) Tmax (maximum temperature of warmest month, Bio5), Twq (mean temperature of warmest quarter, Bio10), Tyr (annual mean temperature, Bio1), Tmin (minimum temperature of coldest month, Bio6) and the precipitation based bioclimatic variables (b) Pyr (annual precipitation, Bio12), Pwq (precipitation of warmest quarter, Bio18) from WorldClim ([17]), as well as two parameters derived from this data source: the continentality index (Conti) from Conrad ([10]), and the Ellenberg quotient (EQ - [15]).
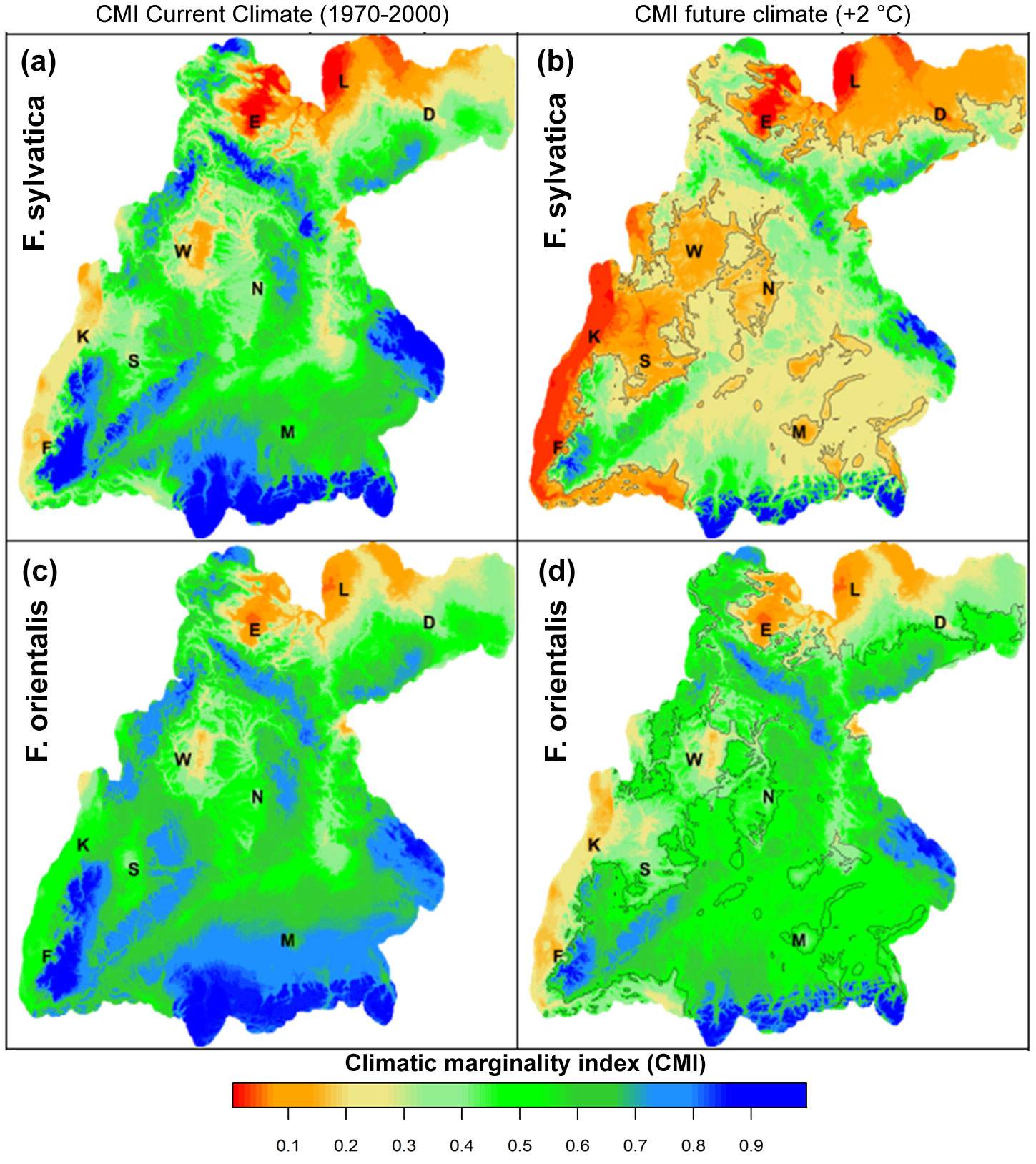
The CMI is more meaningful in climate change studies compared to other habitat suitability indices (HSI - [32]) as it distinguishes between the rear and the leading edge of the climatic range for a species. Recently, it has been used in many studies ([31], [32], [35], [20], [13], [48], [29], [42]). The CMI obtained in this study allowed a subdivision of the species niches into five meaningful zones along the gradient from the rear to the leading edge (see Fig. 3for details): (i) the marginal zone/rear edge (CMI < 0.2); (ii) the intermediate zone/rear edge (CMI = 0.2-0.4); (iii) the optimal zone (center of the distribution, CMI = 0.4-0.6); (iv) intermediate zone/leading edge (CMI = 0.6-0.8); and the marginal zone/leading edge (CMI > 0.8). To highlight the suitability of F. orientalis at marginal F. sylvatica sites in Southern Germany, we added contour lines to the maps delineating the border of the future marginal zone of F. sylvatica (right panels in Fig. 3).
Fig. 3 - Climatic marginality index (CMI) in the study area for F. sylvatica (panels a, b) and F. orientalis (panels c, d) in current (1970-2000; panels a, c) and future climate (+2°C; panels b, d), according to the BIOCLIM algorithm based on the quantile distribution of annual mean temperature (Tyr) and annual precipitation (Pyr) of the whole range of the species distribution ([9]). The contour lines in the right panels (b, d) delineate the prospective marginal area (CMI < 0.2) of F. sylvatica in the future scenario. The position of the main cities is given as black letters. (D): Dresden; (E): Erfurt; (F): Freiburg; (K): Karlsruhe; (L): Leibzig; (M): Munich; (N): Nuremberg; (W): Wurzburg.
For the evaluation of the map outputs, the Pearson’s correlation coefficient between CMI and EQ at points within the target area (n=2048) was applied. The distribution map of European beech by Bohn & Neuhäusl ([3]) containing only the westernmost parts of Oriental beech’s distribution was used as an independent dataset for the evaluation of the map outputs for F. sylvatica.
For all statistical computations and the production of most maps, the packages “raster” and “dismo” ([22]) were used in R ([41]). The map in Fig. 1was produced using the software QGIS ([40]).
Results and discussion
Climatic range
Climatic ranges of both sister species of Fagus overlap each other (Fig. 2). However, temperature and precipitation ranges indicate that F. orientalis reaches warmer and drier climates than F. sylvatica. Temperature ranges of F. orientalis are generally higher than those of F. sylvatica. Twq showed the largest differences with higher values at the median (+1.8 °C), the third quartile (+2.3 °C) and the upper whisker (+4.5 °C). The respective values for Tyr were 1.5 °C, 1.8 °C and 3.4 °C. Tmin showed the smallest range differences, with a mean deviation of +0.8 °C (Fig. 2a).
While temperature ranges of F. orientalis are on a higher thermal level, the species grows under lower precipitation values than F. sylvatica, Especially Pwq values are markedly lower with a median of 156 vs. 224 mm and a lower whisker value of 2 vs. 8 mm compared to F. sylvatica. The respective differences of Pyr values are 738 vs. 787 mm and 215 vs. 362 mm.
Conti interquartiles of both species are within the oceanic/maritime range (20-50) according to the categories of Andrade & Corte-Real ([1]). While lower whisker values of F. sylvatica reach the edge of hyper-oceanic climate, Conti values of F. orientalis remain clearly above this level. Upper whisker values of Conti of both species barely exceed the threshold to a sub-continental climate (Conti = 60). However, extreme occurrences of F. orientalis can be found deeper into a sub-continental climate. Obviously, F. orientalis grows prefentially in a more continental climate, while F. sylvatica prefers a more oceanic climate.
EQ interquartile ranges of F. orientalis from 18.7 to 32.3 show that the core distribution range of this species exceed the aridity limit (EQ = 30 - [15]) of F. sylvatica, while this cannot be observed in records of the latter species. The threshold of an EQ=30 has recently been confirmed by a species distribution study ([32]) and an ecotone investigation ([24]).
A comparison of climatic ranges of both species clearly shows that F. orientalis is well adapted to climate warming (e.g., a Tyr increase of 2 °C) and should better withstand the conditions at marginals sites on the rear edge of F. sylvatica (the third quantile of F. orientalis is 3.4 °C higher). At the same time, F. orientalis tolerates lower minimum temperatures, a wider temperature range (Tmin vs. Tmax - Fig. 2) and a more continental climate (Conti) when compared to F. sylvatica. Furthermore, F. orientalis is quite resistant to frost as supported by the USDA frost hardiness index ([12]), which assigns the same lower limits to both species but a higher upper limit for F. orientalis (8 vs. 7). The late frost resistance of the latter species should also be stronger than that of F. sylvatica, as Muffler et al. ([36]) found that the late frost resistance has a high correlation with the continentality range of a species. Similar climatic properties of both species are reported by Houston Durrant et al. ([25]) and Kandemir & Kaya ([28]). According to Svoboda ([44]) also the saplings of F. orientalis are much more resistant to frost, sun scald and drought stress than those of F. sylvatica. Both frost and drought hardiness are important preconditions for the species suitability at forest sites in SG in the future. Similarly to F. sylvatica, the F. orientalis-complex ([37]) consists of a large variety of genotypes and phenotypes with a high ecological differentiation, especially in growth and bud burst ([28]); for example, a Bulgarian provenance of Fagus orientalis (Gramaticovo) showed a higher sensitivity to frost than F. sylvatica provenances in Southern Germany trials. Proper provenance trials and the selection of the best performing provenances is crucial, especially with respect to the tolerance and adaptation to drought and frost in this species.
Mapping
To check the consistency of the maps obtained, we compared the CMI of F. sylvatica with EQ values and the distribution of forests in Southern Germany according to the natural vegetation distribution ([3]).
The Pearson’s correlation coefficient between CMI and EQ was 0.87 (p < 0.001). Records with a CMI value < 0.2 (marginal to the rear edge) represent points with an EQ > 25, i.e., regions where beech is subdominant (EQ range = 25-30) or even almost absent (EQ > 30, oak forests), according to Ellenberg ([15]).
The standard output of BIOCLIM ([22]) is an ordinary HSI. Whilst HSI values are unimodal and consequently have identical values occuring at the leading and the rear edge (Fig. S1a, Fig. S2 in Supplementary material), CMI increases monotonically towards the leading edge (Fig. S1b). Indeed, the CMI is a measure of the climatic distance of a site to the rear edge. This allows a more intuitive interpretation in terms of climate warming (“the rear edge matters” - [19]). In contrast, in Fig. S2 (Supplementary material), marginal sites (low values) at the rear edge in the lowlands are hardly distinguished from those at the leading edge lying at high altitudes in the south (Alps) and the mountain ranges in the south-west as well as at the border of the middle-east (Fig. 1).
Comparing our results with the map of natural vegetation in Europe ([3]), we found that the areas at the xeric edge represent forest types where European beech is absent or merely scattered (red/orange areas in Fig. 3a). In the large reddish areas at the north-eastern border of Saxony and Thuringia (close to Erfurt and Leipzig) and in the central region in Lower Franconia (close to Wurzburg), the dominant vegetation type is sessile oak-hornbeam forests. In reddish areas in the south-western territory in the lowlands of the river Rhine between Freiburg and Karlsruhe, where European beech is also marginal (red areas), pedunculate oak-hornbeam forests are prevailing ([3]).
The areas at the leading edge (blue colours in Fig. 3a) represent mountain forests with a large proportion of Abies alba, Picea abies and Acer pseudoplatanus. In the Bavarian Alps, the Black Forest and the Bavarian Forest, montane to altimontane types predominate, with an increasing proportion of conifers as site elevation increases.
In summary, the comparison with expert knowledge ([15]) and vegetation maps ([3]) proved that the CMI for our target area for F. sylvatica is quite realistic. We assume that the map produced for F. orientalis should analogously be useful for our purpose, as both maps were developed using the same data set ([9]), data processing procedure and algorithm.
Current suitability of F. sylvatica is high in the studied region, as optimal and intermediate conditions for the species are found in 80% of the study area. The marginal zones at the leading edge (at higher altitudes - blueish colors in Fig. 3a) are much larger (19%) than at the rear edge (2.5% - reddish colors in Fig. 3a). The differentiation of these zones is crucial, as only the sites at the rear edge are under a high risk of local disappearance for F. sylvatica, while sites at the leading edge are largely buffered against climate change ([32]).
Future suitable habitats of F. sylvatica are shown in Fig. 3b. Likely, climate warming by 2 °C will only slightly change the vegetation conditions at higher altitudes (optimal and intermediate CMI). While the marginal areas at the leading edge are reduced from 19% to 7%, the warm and dry zones mostly endangered by warming will be increased about 6-times (15%). For comparison with F. orientalis, the marginal area of F. sylvatica is also outlined by contour lines in the map of F. orientalis (Fig. 3d).
Current suitability of F. orientalis is similar to that of F. sylvatica. In more than 80% of the study area F. orientalis shows at least an intermediate suitability. Also, the marginal areas for F. orientalis are much larger (15%) at the leading edge (at higher altitudes - bluish area in Fig. 3c) than those at the rear edge (1% - red/orange area in Fig. 3a).
Based on the +2 °C warmer scenario, F. orientalis in Southern Germany will remain in optimal/suboptimal conditions in a large area (optimal and intermediate > 80% - Fig. 3d), while the marginal areas at the leading edge is reduced from 15% to 2%, and the mostly endangered zones at the rear edge are small (2%). Interestingly, most parts of the marginal area of F. sylvatica (Fig. 3d) fall within the intermediate/optimal range of F. orientalis. Only the warmest and driest region in the lowlands in the north-east resulted largely unsuitable for F. orientalis.
Our results showed that F. orientalis could be an interesting species with a high potential for future forests in Southern Germany, especially at marginal sites of European beech. However, the approach used in this study has some limitations, such as: (i) the physiological mechanisms of species to face environmental stresses (e.g., drought) was not considered; (ii) the assumption of the consistency of the species ecological niche in the future, as well as the consistency of their future genetic variability and phenotypic plasticity; (iii) the omission of biotic interactions; (iv) the equilibrium assumption; and (v) the disregard of human impact and soil conditions. All the above assumptions were out of the scope of this pilot study and could not be addressed.
In this study, we assumed that current continental-scale patterns of beech occurrence are in equilibrium with climatic patterns, irrespective of the migration history, competitive interactions and human impact ([26]). For a detailed discussion of the limitations of mapping beech distribution, see Mellert et al. ([32]). However, as for species comparison, we used the same data and algorithm for both species, thus avoiding possible bias. According to Hijmans & Graham ([21]), BIOCLIM slightly underestimates range sizes compared to other algorithms (GAM, Domain, Maxent). Further research should include species distribution studies involving inferential statistics based on real observations. Moreover, provenance studies, planting experiments and further investigations on the genetic variability of natural populations should be carried out on a wide range of sites are recommended to further test our findings.
F. orientalis has ecological characteristics quite similar to those of F. sylvatica ([25]), as both species are shade-tolerant, dominant climax tree species with a high growth performance ([28]) and similar ecological niches. Like the European beech, F. orientalis often occurs in a mixture with conifers and could therefore replace F. sylvatica in near-natural woods and forests. This has high practical relevance, as changes in the management form could not be necessary. Moreover, F. orientalis could be used to increase the broadleaves share in conifer forests, thereby converting these stands into mixed close-to-nature forests ([5]) and increasing the ecological and economical advantages of mixed forests ([30]) in submontane and hilly zones of Southern Germany.
The increasing loss of F. sylvatica due to drought events in marginal areas accelerates the transformation of beech to oak forests, partly because of the change in the microclimate of opened stands ([24]). Besides well adapted provenances of F. sylvatica ([34]), the drought-resistant and at the same time shade-tolerant Oriental beech could help to delay this process and keep beech forests on a larger area in the future. Genetic variability and diversity of forest reproductive material will play a key role in forest adaptation under future environmental conditions, and selected seed sources from the natural distribution areas should be used. However, when introducing F. orientalis in Southern Germany, the possible hybridization has to be taken into account, which could at least affect possible local adaptations of the species.
Conclusions
The climatic range of Oriental beech suggest its possible cultivation in critical sites of European beech in German forests, as inferred from simple statistical methods used in this study. Moreover, Oriental beech is interesting to replace European beech in many landscapes in Southern Germany, as its climatic niche is within the current and future climate ranges of the study area. However, before introducing Oriental beech at a large scale, further species distribution studies based on real observations are needed, as well as provenance studies, investigations of the genetic variability of natural populations and field trials in a wide range of sites.
Acknowledgements
This study was funded by the Federal Ministry of Food and Agriculture and the Federal Ministry for the Environment, Nature Conservation, Nuclear Safety and Consumer Protection based on a resolution of the German Bundestag (project number 28W-B-4-058-01) and the Bavarian Office for Forest Genetics. We thank Mrs. Rebekka Stüwe for the linguistic revision of the manuscript.
References
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Online | Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Muhidin Šeho 0000-0001-9926-4564
Bavarian Office for Forest Genetics / Bayerisches Amt für Waldgenetik, Forstamtsplatz 1, 83317 Teisendorf (Germany)
Corresponding author
Paper Info
Citation
Mellert KH, Šeho M (2022). Suitability of Fagus orientalis Lipsky at marginal Fagus sylvatica L. forest sites in Southern Germany. iForest 15: 417-423. - doi: 10.3832/ifor4077-015
Academic Editor
Maurizio Marchi
Paper history
Received: Feb 15, 2022
Accepted: Aug 08, 2022
First online: Oct 19, 2022
Publication Date: Oct 31, 2022
Publication Time: 2.40 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2022
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 29779
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 24246
Abstract Page Views: 2769
PDF Downloads: 2207
Citation/Reference Downloads: 4
XML Downloads: 553
Web Metrics
Days since publication: 1351
Overall contacts: 29779
Avg. contacts per week: 154.30
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2022): 8
Average cites per year: 2.00
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Local ecological niche modelling to provide suitability maps for 27 forest tree species in edge conditions
vol. 13, pp. 230-237 (online: 19 June 2020)
Research Articles
Predicting the effect of climate change on tree species abundance and distribution at a regional scale
vol. 1, pp. 132-139 (online: 27 August 2008)
Review Papers
Impacts of climate change on the establishment, distribution, growth and mortality of Swiss stone pine (Pinus cembra L.)
vol. 3, pp. 82-85 (online: 15 July 2010)
Research Articles
Predicting the impacts of climate change on the distribution of Juniperus excelsa M. Bieb. in the central and eastern Alborz Mountains, Iran
vol. 11, pp. 643-650 (online: 04 October 2018)
Research Articles
The missing part of the past, current, and future distribution model of Quercus ilex L.: the eastern edge
vol. 17, pp. 90-99 (online: 22 March 2024)
Research Articles
Distribution factors of the epiphytic lichen Lobaria pulmonaria (L.) Hoffm. at local and regional spatial scales in the Caucasus: combining species distribution modelling and ecological niche theory
vol. 17, pp. 120-131 (online: 30 April 2024)
Research Articles
Environmental niche and distribution of six deciduous tree species in the Spanish Atlantic region
vol. 8, pp. 214-221 (online: 28 August 2014)
Research Articles
Fluctuation of the ecological niche of Moringa peregrina (Forssk.) Fiori with topoclimatic heterogeneity in southern Iran
vol. 16, pp. 53-61 (online: 16 February 2023)
Research Articles
Reviewing climatic traits for the main forest tree species in Italy
vol. 12, pp. 173-180 (online: 15 March 2019)
Research Articles
Spatial modeling of the ecological niche of Pinus greggii Engelm. (Pinaceae): a species conservation proposal in Mexico under climatic change scenarios
vol. 13, pp. 426-434 (online: 16 September 2020)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword






