The nurse-plant effect under the dislodgement stress of landslides
iForest - Biogeosciences and Forestry, Volume 16, Issue 2, Pages 78-85 (2023)
doi: https://doi.org/10.3832/ifor4017-015
Published: Mar 16, 2023 - Copyright © 2023 SISEF
Research Articles
Abstract
While the mitigating effects of trees on shallow landslide occurrence are well recognised, the impact of landslides on tree community structure and tree-tree interactions have received much less research attention. The structures of tree communities before and after landslides were compared in a 25-ha subtropical forest plot. Tree-tree interactions were examined by analysing the pre- and post-landslide spatial point patterns of large (DBH ≥ 20 cm) and small (1 cm ≤ DBH < 20 cm) tree cohorts. In landslide scarps, 35 (34%) of 104 large trees and 467 (13%) of 3.072 small trees survived. Large (L) and small (S) tree cohorts were paired together for spatial analyses, including pre-landslide (PL) (LPL-SPL), surviving (S) (LS-SS), and missing (M) large-small tree paired cohorts (LM-SM). We randomly selected trees from the pre-landslide tree cohorts to create two virtual paired cohorts, the L34%-S13% and L66%-S87% paired cohorts, whose population sizes were identical to the field-observed LS-SS and LM-SM paired cohorts respectively, but with random spatial patterns. Post-landslide survival rates of trees increased monotonically with DBH. Large trees dislodged by landslides scarcely reduced small-tree survival. Evidence for this included: (i) the distance from small trees to the nearest large trees of the LM-SM paired cohort did not differ significantly from that of the virtual L66%-S87% paired cohort; (ii) survival rates of small trees near LM individuals did not differ significantly from those without large trees nearby. Surviving large trees had positive effects on the survival of small trees, indicated by: (i) the distance from small trees to the nearest large trees of the LS-SS paired cohort was significantly lower than that of the virtual L34%-S13% paired cohort; (ii) SS individuals clumped around LS individuals, whereas the virtual L34%-S13% spatial relationship was random. Large trees prevent landslide dislodgement of adjacent small trees through the nurse-plant effect. Our study suggests that landslide damage in sloping forests may be reduced simply by constantly maintaining a critical density of large trees.
Keywords
Facilitation, Forest Dynamics Plot, Nurse-plant Effect, Point Pattern Analysis, Slope Stability
Introduction
Forests are known to reduce the impacts of shallow landslides through their abiotic mechanical and hydrologic properties ([21], [4], [44]). Shallow landslides in forests typically involve a sliding surface soil mass with a depth of up to 2 m ([41]). The tensile and compressive resistance of tree roots increase the shear strength of shallow soils and, in turn, slope stability ([10]). Forests also reduce the pore water pressure of soils through rainfall interception and transpiration, thereby increasing the intrinsic mechanical strength of soil and the stability of a forested slope ([11], [38]).
Biotic interactions in forests are numerous and wide-ranging ([39]), and can be negative (e.g., competition - [17], [33]) or positive (e.g., facilitation - [22], [33]), and interspecific ([17], [22]) or intraspecific ([17], [33]). Biotic interactions between trees influence tree community properties, including trees’ species diversity ([23]), spatial distribution ([15]), and stem density ([26]). The likelihood of landslide occurrence has been related to tree species diversity ([14], [30]), spatial distribution ([34], [10], [9]), and stem density ([12], [28]). It follows that tree-tree interactions should influence landslide occurrence through their effects on tree community properties. On the other hand, the intensity of plant-plant interactions varies with species diversity, individuals’ spatial distribution, and stem density of plant communities ([39]). Since landslides can substantially change these three properties of tree communities ([44]), landslides are likely to exert effects on tree-tree interactions through altering tree community properties. However, few studies explore the interplay between landslides and tree-tree interactions.
While biotic interactions between trees under post-landslide chronic stresses are well appreciated ([44]), the processes by which the acute stresses of landslide dislodgement influence the post-landslide tree communities and tree-tree interactions are only beginning to be explored. Since landslides cause rapid plant mortality through uprooting and burial ([18]), landslide dislodgement is a form of acute stress for plant survival. Although the loss of tree biomass is amongst the most noticeable landslide effects on forests, the processes by which landslides alter tree community structure are largely unknown because of the lack of pre-landslide vegetation data. Moreover, most previous studies disproportionally focused on how chronic stresses induced in landslide scarps, such as drought or high solar radiation, affect vegetation succession ([44]). Under acute landslide dislodgement stress, tree-tree interactions through roots, such as root grafting, may change tree survival rates. During dislodgement by landslides, the effects of proximate large trees on small tree survival will be negative if small trees in the root zones of dislodged large trees are likely to be removed or buried. Conversely, the biotic interactions between large and small trees will be positive if small trees in the root zones of surviving large trees are kept in place. Identifying the transition of tree community structure and tree-tree interactions associated with landslide dislodgement could provide new insights into landslide-prone forest management.
The present study aimed to identify landslide impacts on tree community structure and interactions between large and small trees under landslide dislodgement stress and to explore the role of such biotic interactions in slope stability. We asked: (i) does the diameter at breast height (DBH) distribution of trees surviving landslides differ from that of the pre-landslide tree communities; (ii) do landslides change the spatial relationships between large and small trees; (iii) is the role played by large trees negative or positive in the survival of small trees?
Materials and methods
Study area
The study site was located in the 25-ha Lienhuachih Forest Dynamics Plot (500 × 500 m), central Taiwan (23° 54′ 49″ N, 120° 52′ 43″ E). The elevation ranges from 667 m to 845 m a.s.l. (Fig. S1 in Supplementary material). This plot is characterised by steep terrain, with an average slope of 36° and a maximum of 77°. The annual precipitation is 2439 mm, with most falling in five months (May to September - Fig. S2). The mean annual temperature is 20.4 °C, with mean monthly temperatures ranging from 14.4 °C in January to 24.5 °C in July (Fig. S2 in Supplementary material).
The first tree census for this plot was completed in early 2008. Every tree with a diameter at breast height (1.3 m, DBH) equal to or greater than 1 cm was surveyed. The DBH, species identity, and location of 153.261 trees were recorded. This evergreen broad-leaved forest comprised 144 tree species from 46 families, dominated by Fagaceae, Lauraceae and Rubiaceae ([5]). The overall DBH distribution of tree species was negative exponential, indicating a high proportion of seedlings and saplings (Fig. S3 in Supplementary material). The maximum DBH was 115 cm, and trees with DBHs smaller than 20 cm accounted for 96% of the total stem number. The average overstorey canopy height measured by the airborne LiDAR was 11.4 m ([8]).
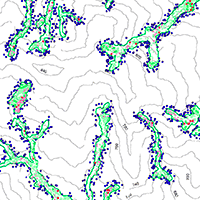
In 2008, typhoons Kalmaegi (maximum rainfall intensity, 450 mm/24 hr; date influencing Taiwan, 17-18 July) and Sinlaku (520.5 mm/24 hr; 14-15 September) caused 11 landslides in our study site ([6]). Most landslide scarps were relatively long and narrow, aligned along valleys (Fig. 1). Using the landslide classification system of Varnes ([42]), the movement type was debris slide (LW Chang, unpublished data). The total disturbed area was 9159.47 m2, and the areas of the largest and smallest landslides were 2561.62 m2 (landslide A) and 83.85 m2 (landslide B), respectively (Fig. 1). Landslides B and J were excluded from the following analyses due to their small areas (Fig. 1).
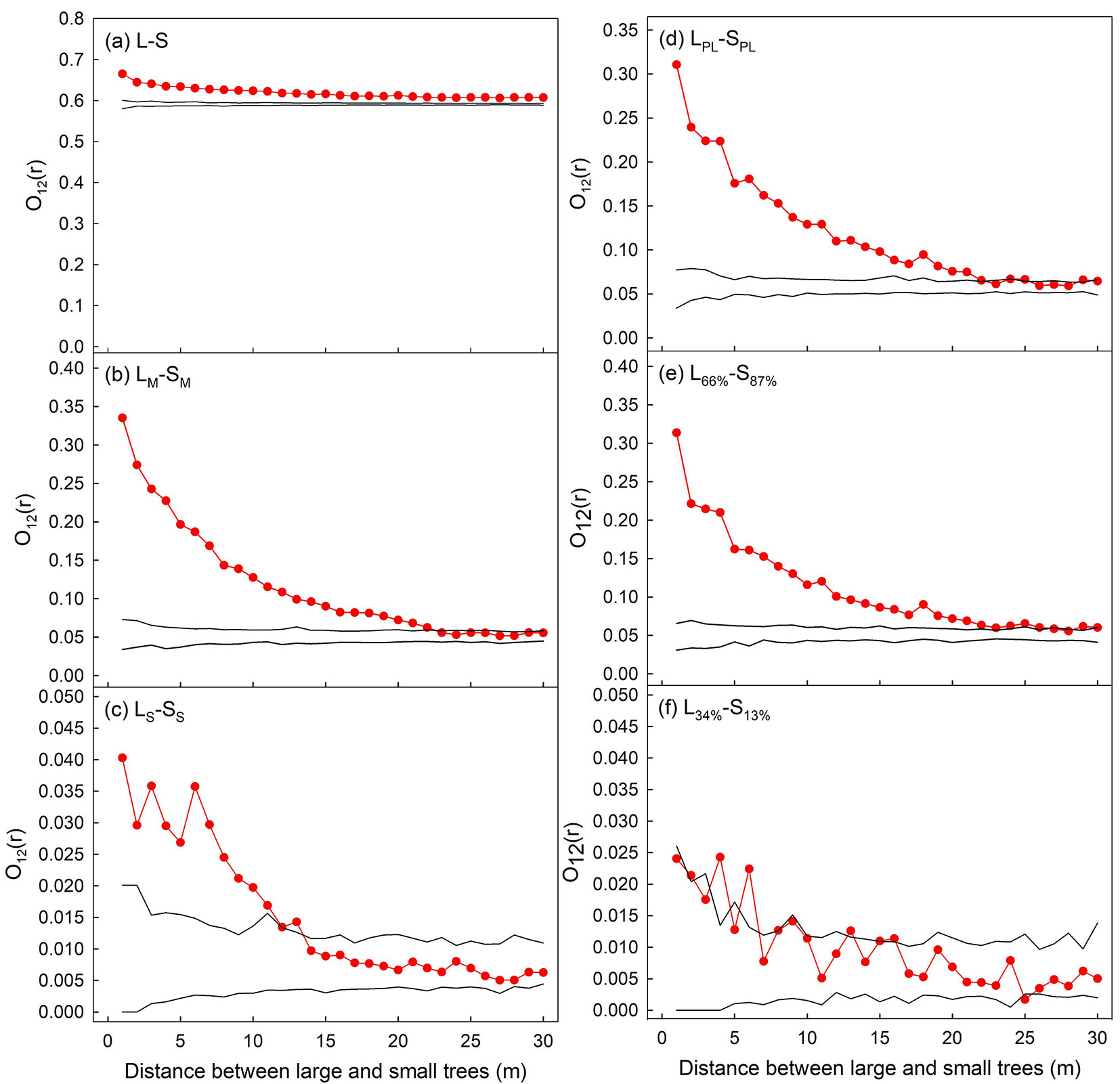
Fig. 1 - The spatial distribution of (a) the 11 landslides and (b) individuals of LS, LM, SS, SM, and LA. These landslides were caused by heavy rains of typhoons Kalmaegi and Sinlaku in 2008. LS, LM, SS, SM, and LA are the abbreviations for the cohorts of surviving large trees, missing large trees, surviving small trees, missing small trees, and large trees around the landslide scarps, respectively. Note that Landslides B and J were not included in our analyses due to their small area.
Data analysis
The depth of landslides was spatially heterogeneous in our study site (LW Chang, unpublished data). It is impossible to choose a threshold DBH with which we could precisely divide trees into large (the protectors against landslide dislodgement stress) and small trees (the protectees) because the threshold DBH should vary with landslide depth. Nevertheless, we selected 20 cm as the threshold DBH with the following approaches. If the threshold DBH is too low, many protectees will be misidentified as protectors. We rejected threshold DBHs of 30 cm and 40 cm, which yielded too few large trees to conduct statistically meaningful analyses. At 20 cm DBH, the 18 most dominant tree species in our study site reached the average overstorey canopy height of 11.4 m (GZM Song, unpublished data). Using the threshold DBH of 20 cm not only minimised the two analytical problems mentioned above but also has strong significance for its ecological (e.g., light interception) and hydrological effects (e.g., rainfall interception) of overstorey trees in our analyses.
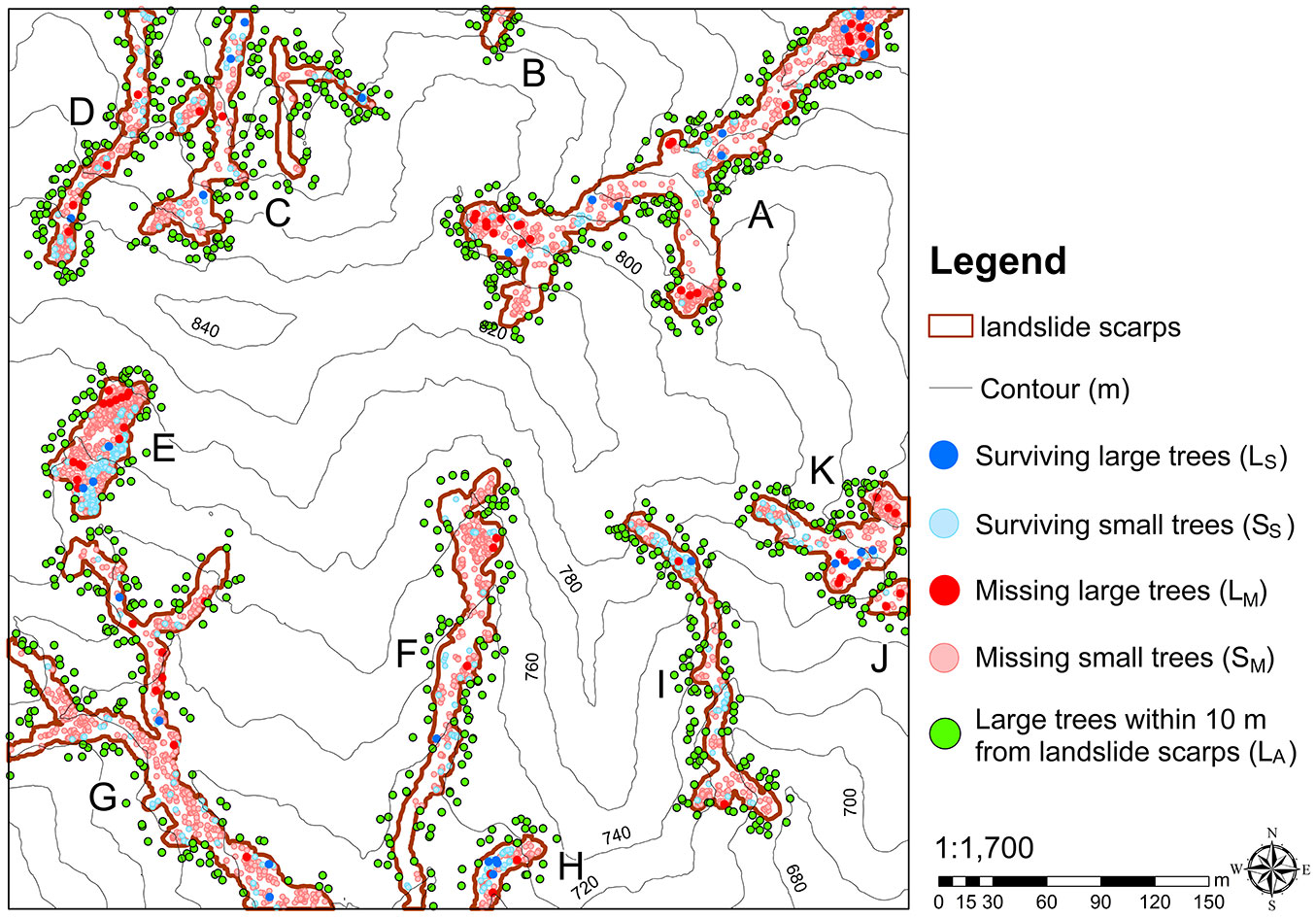
The tree census conducted before the 2008 landslide event recorded 5.449 large trees (L cohort) and 147.812 small trees (S cohort) in the 25-ha plot. The pre-landslide population within landslide scarps created in 2008 was 3.643 trees, including 104 large trees (LPL cohort, L for large tree and PL for pre-landslide) and 3.539 small trees (SPL cohort, S for small tree - see Tab. S1 in Supplementary material). In addition, 934 trees (LA cohort, A for around) were located less than 10 m away from the landscape scarps (Fig. 1, Tab. S1). Field surveys in 2009 showed that, after the 2008 landslide events, 35 large trees (LS cohort, S for surviving) and 467 small trees (SM cohort, M for missing) survived (Fig. S4, Tab. S1), which accounted for 34% and 13% of the pre-landslide populations, respectively ([6]). As a corollary, 66% of large trees and 87% of small trees were missing after the landslides.
To identify spatial relationships affected by biotic interactions between trees, four virtual tree cohorts were created through randomly selecting trees from the pre-landslide tree populations. The population sizes of the four virtual cohorts (L34%, L66%, S13%, and S87%) were identical to those of the LS, LM, SS, and SM cohorts, respectively. Using the L34% cohort as an example, random selection was conducted through allocating a random number to each pre-landslide large tree (LPL) and selecting those trees with numbers from the 1st to the 34th percentile ranks to create this virtual cohort, regardless of their location within the landslide area. The field-observed spatial patterns were removed from the virtual cohorts through random selection. As the population size of the virtual cohorts was controlled, inconsistent results of spatial analyses between the virtual and field-observed cohorts could only be attributed to their different spatial patterns, allowing us to identify trees’ field-observed spatial patterns influenced by tree-tree interactions. In spatial analyses, large-tree cohorts were paired with small-tree cohorts in the same circumstances, e.g., the cohort of missing large trees was paired with that of missing small trees (LM-SM paired cohort).
We located every small tree in the LPL-SPL, LM-SM, virtual L66%-S87%, LS-SS, and virtual L34%-S13% paired cohorts first, and then the nearest large tree for each small tree with the computer program ArcGIS® v. 10.2 (ESRI, Redlands, California, USA). Ultimately, the distances from small trees to the nearest large trees for these paired cohorts were identified. Since the data of some tree cohorts were not normally distributed, differences between groups were always examined with the Mann-Whitney test (a nonparametric method) with Bonferroni correction (a method to counteract inflated Type I errors caused by multiple comparisons).
We used the O-ring statistic, one of the point-centred statistical methods, to explore the spatial relationships between large and small trees. The O-ring statistic is widely used in spatial distribution studies for organisms ([40]) and landslides ([48], [32]). The O-ring statistic includes univariate and bivariate analyses; the former estimates the spatial relationships of individuals in the same group, and the latter examines the spatial correlations between individuals of two groups (Pattern 1 and Pattern 2 - [45], 2014). The present study used bivariate analyses, and large and small tree cohorts were treated as Pattern 1 and Pattern 2, respectively. The O12 estimate in bivariate analyses was calculated as the number of small tree individuals at a specific distance from the centre of the ring (a large tree) ([45], [46]). The equation is (eqn. 1):
where Riw(r) is the ring centred in the ith large tree, with radius r and width w; Points2[Riw(r)] is the total point number of small trees in the ring Riw(r); Area[Riw(r)] is the area of the ring. If a value of O12 at a specific distance from a large tree is greater than the upper limit of the confidence belt, it indicates that small trees are clumped around large trees. Conversely, if the value of O12 is less than the lower limit of the confidence belt, it indicates a repulsion pattern between large and small trees. A value of O12 within the confidence belt represents a random spatial relationship between large and small trees. The O-ring analysis was conducted using the Programita software ([46]). The WM edge correction developed by Wiegand & Moloney ([45], [46]) was used to correct the edge effect.
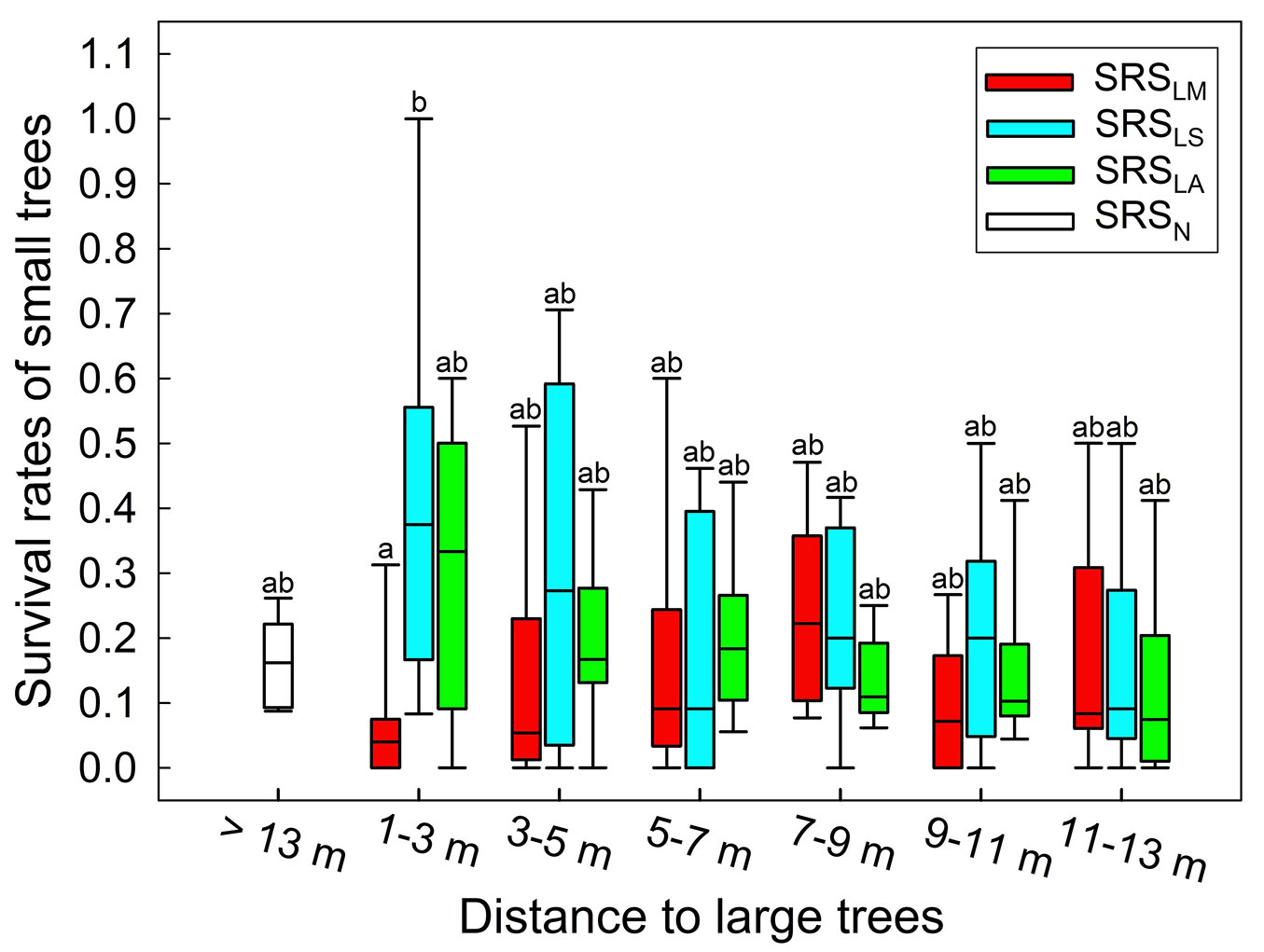
The preliminary O-ring statistic showed that the clumped patterns of SS trees were observed up to 13 m from LS trees. Therefore, this distance was regarded as the maximum distance that the large-tree effects could reach in the survival rate analyses of small trees (SRSs). In the nine landslide scarps, the survival rate for small trees (SRSs) at selected distances (1-3 m, 3-5 m, 5-7 m, 7-9 m, 9-11 m, and 11-13 m) from LS, LM, and LA individuals were analysed, namely SRSLS, SRSLM, and SRSLA, respectively. Survival rates of small trees more than 13 m away from LS, LM, and LA (SRSN) were treated as small-tree survival in the absence of large-tree effects. The Mann-Whitney test (a nonparametric method) with Bonferroni correction was used to identify differences between these small-tree survival rates because of the non-normal distributions of small-tree survival rates.
Results
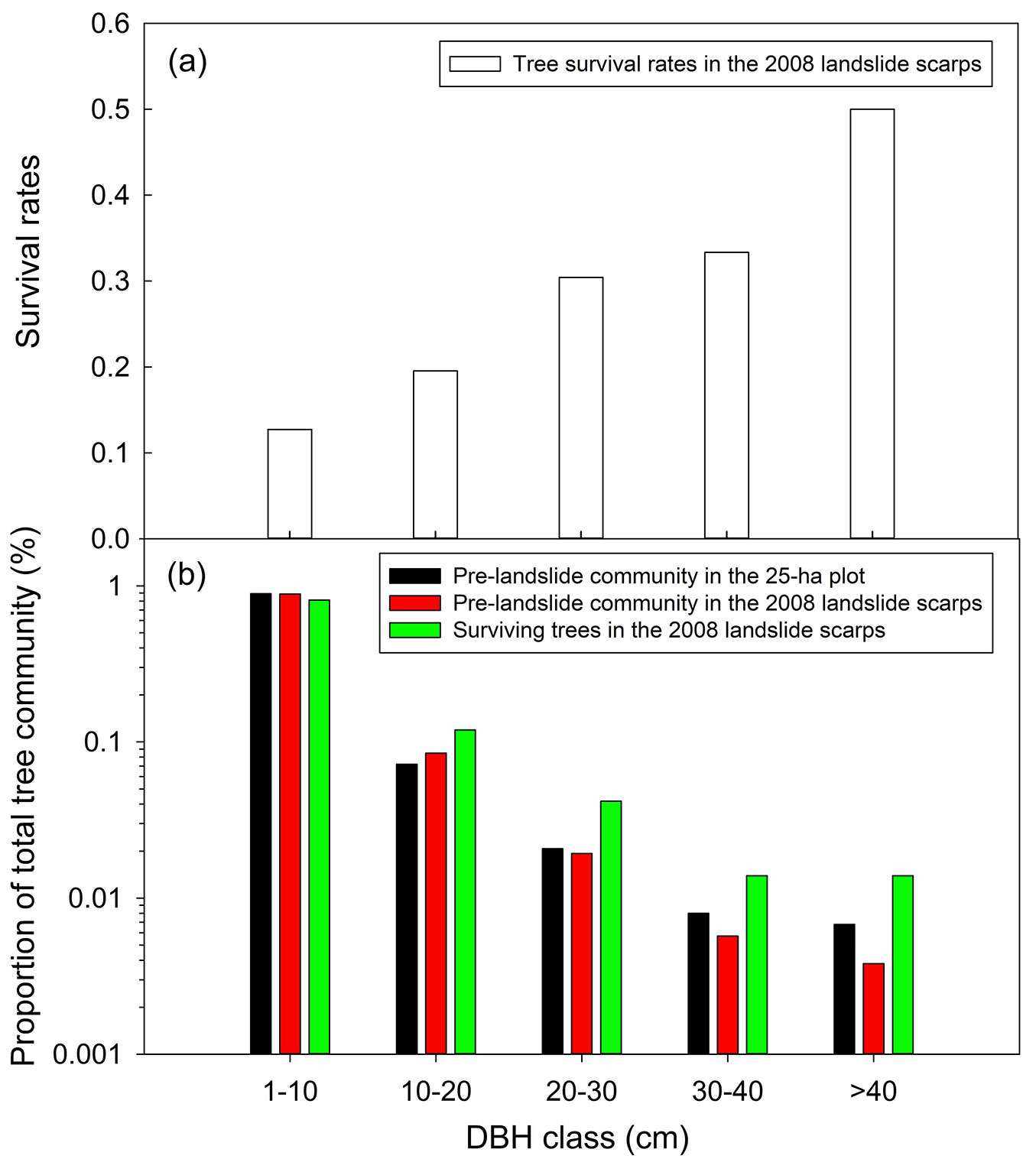
The DBH of surviving trees in the 2008 landslide scarps was significantly larger than that of missing trees and trees outside landslide scarps (p < 0.01, Mann-Whitney test with Bonferroni correction - Fig. 2). Moreover, tree survival rates increased monotonically with DBH (Fig. 3a), so the post-landslide communities had fewer smaller trees compared with the pre-landslide tree communities in the 25-ha plot and in the 2008 landslide scarps (Fig. 3b).
Fig. 2 - DBH of trees outside landslide scarps, missing and surviving trees in landslide scarps. Each bar and error bar respectively represent the mean and standard error of DBH in the same group. Different letters next to bars indicate significant differences (p < 0.05) between groups.

Fig. 3 - Landslide impacts on tree community structure. (a) Tree survival rates associated with DBH class in landslide scarps, and (b) the DBH distributions of the pre- and post-landslide tree communities. Each bar in panel (a) represents the overall tree survival rate of the nine landslide scarps for a DBH class.
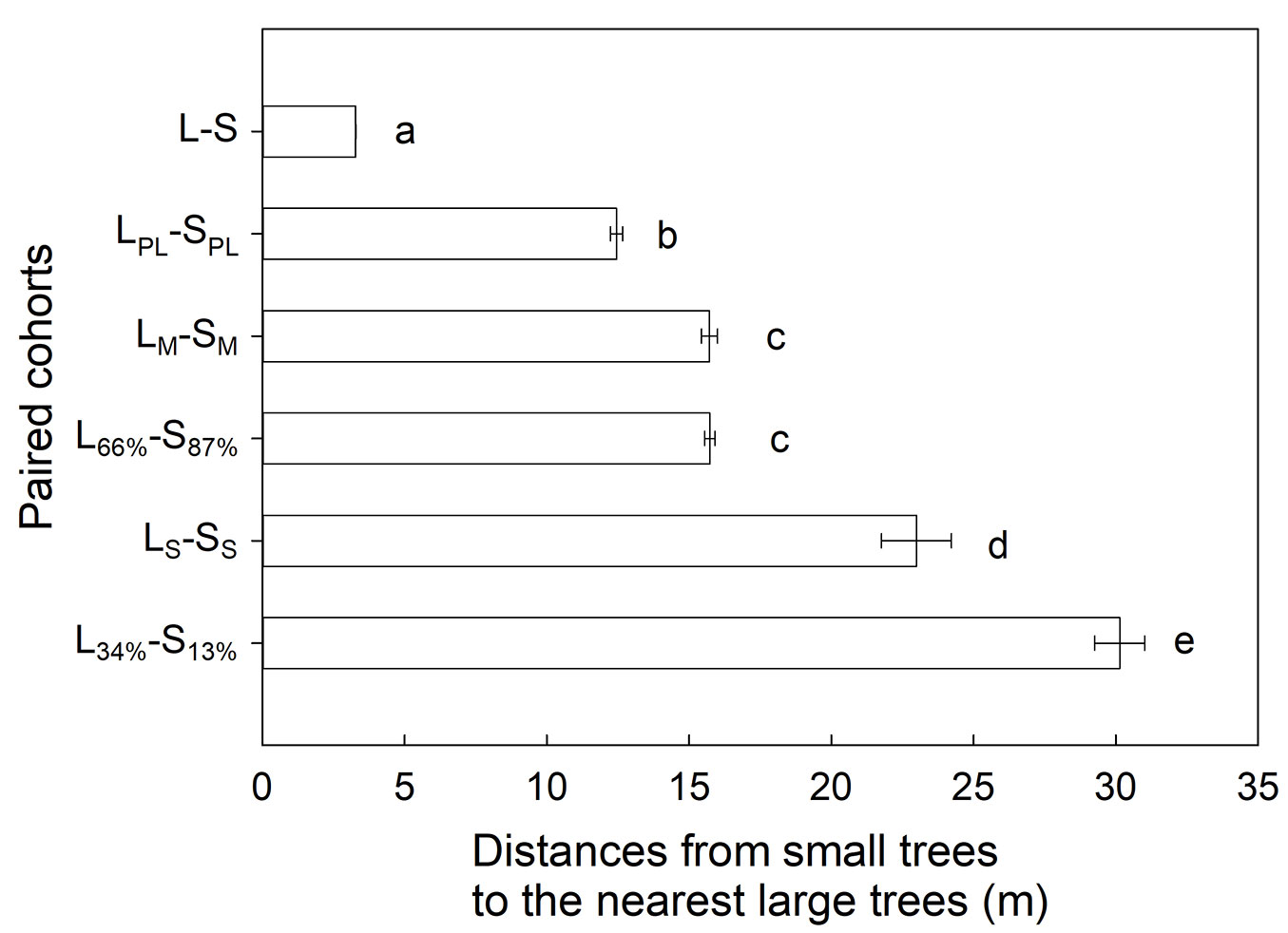
For the five paired cohorts (LPL-SPL, LM-SM, virtual L66%-S87%, LS-SS, and virtual L34%-S13%), the distances from small trees to the nearest large trees generally increased as their population size decreased (Fig. 4). The mean distance from SPL individuals to the nearest LPL individuals was 12.5 m. After the landslides, there were more missing trees (LM, SM) than surviving trees (LS, SS - Fig. 1), so that the distances from SM individuals to the nearest LM were lower than the distances from SS individuals to the nearest LS (Fig. 4). For the more numerous field-observed LM-SM and virtual L66%-S87% paired cohorts, there was no significant difference between the two paired-cohorts (p > 0.05, Mann-Whitney test with Bonferroni correction). In contrast, for the less numerous field-observed LS-SS and virtual L34%-S13% paired cohorts, the distances from SS to the nearest LS were lower than those from S13% individuals to the nearest L34% individuals (p < 0.01, Mann-Whitney test with Bonferroni correction - Fig. 4).
Fig. 4 - The distances from small trees to the nearest large trees for the five paired cohorts. Each bar and error bar respectively represent the mean and standard error of the distances from small trees to the nearest large trees in the nine landslide scarps. Different letters next to bars indicate significant differences (p < 0.01, Mann-Whitney test with Bonferroni correction) between groups. Please refer to the list of abbreviations for the definition of symbols.
Among the six paired cohorts, the virtual L34%-S13% paired cohort was the only one exhibiting a mostly random spatial pattern, while the other five showed clumped patterns (Fig. 5). Comparing the spatial patterns for those paired cohorts with identical population sizes, the LM-SM and the virtual L66%-S87% paired cohorts were clumped (Fig. 5b, 5e), while the LS-SS and virtual L34%-S13% paired cohorts exhibited different patterns (Fig. 5c, 5f). In contrast to the mostly random pattern of the virtual L34%-S13% paired cohort, the clumped pattern of LS-SS paired cohorts indicated the mechanical supports of LS individuals to SS individuals under the dislodgement stress of landslides.
Fig. 5 - Spatial patterns of the five paired cohorts in Lienhuachih Forest Dynamics Plot: (a) L-S, (b) LM-SM, (c) LS-SS, (d) LPL-SPL, (e) virtual L66%-S87%, and (f) virtual L34%-S13% paired cohorts. Two black curves represent the upper and lower borders of the 95% confidence belt. Red curves above and within confidence belts, respectively, indicate clumped and random distribution patterns between paired cohorts. Please refer to the list of abbreviations for the definition of symbols.
Although survival rates of small trees tended to increase with their distance from LM individuals and decrease with their distance from LS and LA individuals, survival rates between groups did not differ significantly (p > 0.05, Mann-Whitney test with Bonferroni correction), except for those less than 3 m away from LS and LM trees (Fig. 6). SRSLA and SRSLM did not differ significantly from SRSN (Fig. 6), indicating that the effects of LA and LM trees on the survival of small trees in landslide scarps were negligible.
Fig. 6 - Survival rates of small trees (SRSs) at different distances from LM, LS, and LA individuals. SRSN represents the survival rates of small trees without any large trees nearby. Different letters above boxes indicate significant differences (p < 0.05) between groups.
Discussion
The role of large trees in the survival of small trees
The spatial patterns of the LS-SS paired cohorts were mainly determined by the nurse-plant effect, and the negative effects of large trees on small tree survival were trivial. In general, the distances from small trees to the nearest large trees increased as the tree population sizes decreased (Fig. 4). Although the population sizes of the field-observed LS-SS and virtual L34%-S13% paired cohorts were identical, the distances from SS individuals to the nearest LS individuals were significantly shorter than those from S13% individuals to the nearest L34% individuals. This phenomenon is attributed to the nurse-plant effect with which LS saved small trees nearby from landslide dislodgment. On the other hand, the distances from SM individuals to the nearest LM individuals should be significantly shorter than those from S87% individuals to the nearest L66% individuals if small trees near LM individuals tend to be dislodged. However, this was not the case (Fig. 4), indicating that the negative effect of missing large trees on small-tree survival was negligible.
The clumped spatial patterns of the LM-SM paired cohorts were mainly determined by the pre-landslide spatial patterns, and the effects of large trees on small tree survival were mainly positive. Post-landslide cohorts (LM, LS, SM, and SS) were sub-populations of the pre-landslide cohorts (LPL and SPL), so that the legacy of the pre-landslide spatial patterns should still be evident after the disturbance. The O-ring statistic showed that, prior to landslides, small trees clumped around large trees (Fig. 5a, Fig. 5d). Such a spatial pattern between small and large trees could result from intraspecific interactions (e.g., the highest seed-fall density near the parent trees -[20]), interspecific interactions (e.g., beneficial mycorrhizal associations - [37]), or abiotic effects (e.g., an uneven distribution pattern of the light environment - [27], favourable soil conditions). For the two virtual paired cohorts, the L66%-S87% paired cohort exhibited a clumped spatial pattern similar to that of the paired LPL-SPL cohort, while the spatial pattern of the L34%-S13% paired cohort was mostly random (Fig. 5d, Fig. 5e, Fig. 5f). These results showed a trend that the spatial correlation between the randomly selected large and small trees changed from clumped to random as the population size of the virtual cohorts decreased. In other words, random selection can remove the legacy of the pre-landslide spatial patterns if the population size of the virtual cohorts is reduced substantially. Therefore, the spatial pattern of the field-observed LM-SM paired cohort, which shared identical population size and similar spatial patterns with the virtual L66%-S87% paired cohorts, should be attributed to the legacy effect (Fig. 5b, Fig. 5e). Since the virtual L34%-S13% paired cohort exhibited a mostly random pattern, the clumped spatial pattern of the field-observed Ls-Ss paired cohort, which population size was identical to the L34%-S13% paired cohort, should be attributed to a nurse-plant effect rather than reflecting the initial spatial pattern of the forest, or the legacy effect (Fig. 5c, Fig. 5f). In summary, under the dislodgement stress of landslides, the role of large trees in small tree survival was mainly positive.
Survival rate analyses also showed that large trees dislodged by landslides had negligible negative effects on small tree survival. Although the survival rates of small trees located 1-3 m away from LM (SRSLM) were significantly lower than those of small trees 1-3 m away from LS (SRSLS), the survival rates of small trees near LM individuals (SRSLM) were not significantly lower than that for small trees without large trees nearby (SRSN - Fig. 6).
Mechanisms of the nurse-plant effect under landslide dislodgement stress
Large tree attributes associated with the survival of small trees after landslide dislodgement should vary with the inter-tree distance. Small trees within or at the edge of large-tree root zones, plus the soils in which they grow, are protected by direct mechanical support and anchorage of large-tree roots. Root grafting, which is the natural fusion of roots of different individuals ([24]), can increase the anchorage of trees in the root network and, in turn, raise their resistance to uprooting caused by winds, floods or landslides ([1], [24]). Although further studies are needed to identify the contribution of root grafting to the nurse-plant effect, crossing over of roots of different individuals is commonly observed in our root excavations for root pullout tests (GZM Song, personal observation), whereby large trees can provide direct mechanical support to small trees.
The protection zones for small trees of LS individuals could be as wide as 13 m (Fig. 5c), which was much greater than the field-observed radii of tree root zones in our study site (GZM Song, personal observation). Within single landslides, SS individuals were significantly closer to the landslide heads than LS individuals (p < 0.01, Mann-Whitney test - Fig. S5 in Supplementary material), suggesting that buttressing and arching of LS roots may have protected small trees even beyond the limits of their root zones. Due to the slope toe protection provided by trees, a location directly upslope from a large tree may be stabilised even though its roots do not reach that point ([10]). This is the so-called buttressing effect. If the distances between trees at hillslope toes are short enough, soil masses with no tree directly down the slope are still stabilised by the root arching effect ([12]). Similarly, forests on the path of debris flow reduce debris-flow runout and, in turn, protect downslope trees or infrastructure ([2]). Since most landslides in our study site occurred in valleys or close to drainage lines (Fig. 1), surviving large trees may have intercepted debris flow and, in turn, reduced the extent of mechanical damage to small trees. Further studies are needed to verify the roles of these effects in the tree-tree interactions under the dislodgement stress of landslides.
Landslide impacts on tree community structure
Landslides change tree community structure by removing more trees of smaller DBHs. Post-landslide tree survival is a result of the interplay between landslide depth and root depth. Tree roots must reach soils deeper than those displaced by landslides if they are to survive landslides. The mean depth of landslides in our study site was 1.25 m (LW Chang, unpublished data), which would allow deeper-rooted trees to survive. Root depth tends to increase with the DBH of trees ([13]). Therefore, in the present case, the DBHs of surviving trees were higher than those of missing trees (Fig. 2), and tree survival rates increased with their DBHs (Fig. 3a). Although landslides lowered the proportion of stem number in small DBH classes, the post-landslide tree community structure maintained a negative exponential DBH distribution (Fig. 3b). Such a community structure was partly a result of the nurse-plant effect with which surviving large trees reduced small tree mortality caused by landslides. The negative exponential DBH distribution is an indication of sustainable tree populations and communities ([36]). In addition, these landslide scarps were surrounded by forests, so tree recruitment could be promoted by seed rains from neighbouring trees. It is expected that the tree community structure in landslide scarps in the study area will remain negative exponential in the future.
Suggestions for landslide-prone forest management and future studies
The nurse-plant effect of large trees should be incorporated into the management of landslide-prone forests. Due to the facts that root reinforcement is mainly contributed by large roots ([16]) and larger trees develop more large roots than do small trees, large trees survived landslides better than did small trees (Fig. 2, Fig. 3). Surviving large trees protected small trees from landslide dislodgement (Fig. 4, Fig. 5) so that soils occupied by these small trees were kept in place. Although root reinforcement from the remaining roots of logged trees will not vanish until a few years later, root decomposition following clearcutting practices may increase landslide risks for several years ([3], [43]). Silvicultural practices associated with retaining large trees, such as the systems of single tree selection, seed tree, or shelterwood ([31]), are recommended to reduce soil loss due to landslides. Results of the present study (Fig. 6), Cislaghi et al. ([9]) and Moos et al. ([28]) indicated that, to maximise the protective effects of large trees, their spacing distance should be kept below 6 m (i.e., average stem density ≥ 256 stem ha-1).
In addition to the acute dislodgement stress of landslides, large trees surviving landslides can ease the chronic post-landslide stresses and, in turn, shorten the time of vegetation recovery. Post-landslide habitat degradation poses several chronic stresses for plant colonisation, such as excessive solar radiation and soil erosion associated with vegetation removal and soil nutrients loss due to topsoil removal ([47], [25], [44], [6]). The long-term colonisation of persistent pioneer plants can further retard the establishment of late-successional species and, in turn, arrest vegetation succession ([18], [29], [35]). Through intercepting strong sunlight, providing materials for soil organic matter enrichment, and suppressing the growth of persistent pioneer plants, large trees in or near landslide scarps can exert positive effects on the growth and survival of recruited and existing small trees and ultimately promote vegetation recovery ([44], [7]).
Improved management of forests susceptible to landslides requires more studies to unveil the interplay between tree-tree biotic interactions and landslides. Our results showed that the nurse plant effect reduces small tree mortality in landslides. Therefore, increasing or at least maintaining a critical stem density of large trees can be employed as both preventative (reducing tree loss to landslides) and restorative strategies (easing post-landslide drought and harsh sunlight effects) for hillslope forest management. On the other hand, broader biotic interactions may also influence landslide phenomena. For example, allelopathy, mycorrhizal systems, and the Janzen-Connell effect may change tree community properties (e.g., root density, spatial distribution, stem density - [15], [37], [19]) and, in turn, the likelihood and impact of landslide occurrence. Further detailed studies linking biotic interactions to landslide occurrence would enable biotic interactions to be incorporated into management tools for landslide-prone forests.
Conclusion
The present study aimed to examine how shallow landslides influenced the structure and tree-tree interactions of tree communities. Although landslide survival increased with increased tree DBH, the DBH distribution of the post-landslide tree community structure retained the negative exponential form of the pre-landslide forest. Landslide impacts on tree regeneration were expected to be trivial due to this continuity of community structure and the numerous trees surrounding landslide scarps. The nurse-plant effect of large trees on small tree survival in landslide scarps was identified by two phenomena: the clumping of surviving small trees around surviving large trees, and the significantly smaller mean distance between members of surviving large and small tree paired cohorts compared with the corresponding virtual paired cohort. The survival rates of small trees within 3-13 m from missing large trees were not significantly lower than those with no large trees nearby, indicating that large trees did not increase the mortality of adjacent small trees once they were dislodged. Our study established that interactions between large and small trees under the dislodgement stress of landslides were mainly positive and that such biotic interactions can reduce tree mortality resulting from landslides. We suggest that hillslope forests susceptible to shallow landslides should be managed through the simple procedure of maintaining a specified minimum density of large trees and recommend further studies into the relationships between biotic interactions and landslides.
List of abbreviations
(L): the cohort of large trees recorded in the 25-ha plot before the 2008 landslide event; (L34%, L66%): cohorts consist of trees randomly drawn from LPL. The percentage indicates their population size related to that of the LPL cohort; (LS): the cohort of surviving large trees in the 2008 landslide scarps; (LM): the cohort of large trees removed by the 2008 landslides; (LA): the cohort of large trees less than 10 m away from the 2008 landslide scarps; (LPL): the cohort of pre-landslide large trees in the 2008 landslide scarps; (S): the cohort of small trees recorded in the 25-ha plot before the 2008 landslide event; (S13%, S87%): cohorts consist of trees randomly drawn from SPL. The percentage indicates their population size related to that of the SPL cohort; (SS): the cohort of surviving small trees in the landslide scarps; (SM): the cohort of small trees removed by the 2008 landslides; (SPL): the cohort of pre-landslide small trees in the 2008 landslide scarps; (SRSN): survival rates of small trees which were at least 13 m away from LS, LM, and LA; (SRSLS): survival rates of small trees near LS; (SRSLM): survival rates of small trees near LM; (SRSLA): survival rates of small trees near LA.
Acknowledgements
This study was funded by the Taiwan Ministry of Science and Technology (MOST 108-2625-M-005-007), the Taiwan Forestry Research Institute (97 AS-7.1.1.F1-G1), and the Forestry Bureau of Taiwan (TFBM-960226). We would like to express our gratitude to the crews of the Lianhuachi Research Center of the Taiwan Forestry Research Institute for field data collection and data entry. We thank Drs. Dar-Hsiung Wang, Han-Ching Hsieh, Yu-Jen Chiang, Pai-Han Yang, Tsai-Huei Chen, Chin-Shien Wu for providing the digital terrain model of our study site.
Authors Contribution
Jian-Hong Yang: Conceptualisation, Data Curation, Formal analysis, Methodology, Visualisation, Writing - Original Draft, Writing - Review & Editing. Li-Wan Chang: Data Curation, Funding acquisition, Investigation. Kai-Chi Hsu: Project administration, Writing - Review & Editing. Chia-Cheng Fan: Validation, Writing - Review & Editing. David Doley: Writing - Review & Editing. Guo-Zhang M. Song: Conceptualisation, Funding acquisition, Methodology, Project administration, Supervision, Visualisation, Writing - Original Draft, Writing - Review & Editing.
References
Online | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Kai-Chi Hsu
Guo-Zhang Michael Song 0000-0002-9266-3690
Department of Soil and Water Conservation, National Chung Hsing University, 145 Xingda Rd., South Dist., Taichung City 402 (Taiwan)
Botanical Garden Division, Taiwan Forestry Research Institute, No.67 Sanyuan St., Zhongzheng Dist., Taipei City 100051 (Taiwan)
Forest Protection Division, Taiwan Forestry Research Institute, No.67 Sanyuan St., Zhongzheng Dist., Taipei City 100051 (Taiwan)
Department of Construction Engineering, National Kaohsiung University of Science and Technology, No.1, University Rd., Yanchao Dist., Kaohsiung City 824 (Taiwan)
Centre for Mined Land Rehabilitation, The University of Queensland, Brisbane, Qld 4072 (Australia)
Corresponding author
Paper Info
Citation
Yang J-H, Chang L-W, Hsu K-C, Fan C-C, Doley D, Song G-ZM (2023). The nurse-plant effect under the dislodgement stress of landslides. iForest 16: 78-85. - doi: 10.3832/ifor4017-015
Academic Editor
Giorgio Vacchiano
Paper history
Received: Nov 11, 2021
Accepted: Dec 18, 2022
First online: Mar 16, 2023
Publication Date: Apr 30, 2023
Publication Time: 2.93 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2023
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 24734
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 20082
Abstract Page Views: 2401
PDF Downloads: 1783
Citation/Reference Downloads: 1
XML Downloads: 467
Web Metrics
Days since publication: 1198
Overall contacts: 24734
Avg. contacts per week: 144.52
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2023): 1
Average cites per year: 0.33
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Discovering interaction between oaks and carabid beetles on a local scale by point pattern analysis
vol. 9, pp. 618-625 (online: 06 May 2016)
Research Articles
Scale dependency of the effects of landscape structure and stand age on species richness and aboveground biomass of tropical dry forests
vol. 16, pp. 234-242 (online: 23 August 2023)
Research Articles
Differences in early dynamics and effects of slope aspect between naturally regenerated and planted Pinus sylvestris woodland on inland dunes in Poland
vol. 9, pp. 875-882 (online: 17 June 2016)
Research Articles
Woody species recruitment under monospecific plantations of pioneer trees - facilitation or inhibition?
vol. 5, pp. 1-5 (online: 06 February 2012)
Research Articles
Monitoring spatial and temporal pattern of Paneveggio forest (northern Italy) from 1859 to 2006
vol. 3, pp. 72-80 (online: 17 May 2010)
Research Articles
Slope shape effect on runoff and soil erosion under natural rainfall conditions
vol. 7, pp. 110-114 (online: 18 December 2013)
Review Papers
Large-scale effects of forest management in Mediterranean landscapes of Europe
vol. 6, pp. 342-346 (online: 29 August 2013)
Research Articles
The effect of the calculation method, plot size, and stand density on the accuracy of top height estimation in Norway spruce stands
vol. 10, pp. 498-505 (online: 12 April 2017)
Research Articles
Historical fire ecology and its effect on vegetation dynamics of the Lagunas de Montebello National Park, Chiapas, México
vol. 14, pp. 548-559 (online: 01 December 2021)
Research Articles
Evaluation of urban forest landscape health: a case study of the Nanguo Peach Garden, China
vol. 13, pp. 175-184 (online: 02 May 2020)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword






