Biodiversity conservation and wood production in a Natura 2000 Mediterranean forest. A trade-off evaluation focused on the occurrence of microhabitats
iForest - Biogeosciences and Forestry, Volume 12, Issue 1, Pages 76-84 (2019)
doi: https://doi.org/10.3832/ifor2617-011
Published: Jan 24, 2019 - Copyright © 2019 SISEF
Research Articles
Abstract
The most significant European forest-related strategies highlight the importance of multifunctional forests for human wellbeing, due to the provision of a wide range of goods and services. However, managing competing aims, such as timber production, economic drivers and biodiversity conservation is often difficult for practitioners. In order to assess the loss and gain of ecosystem services caused by forestry, trade-off evaluation has been increasingly used to aid decision-making. In this study, four silvicultural scenarios are simulated using the Marteloscope approach to evaluate the trade-offs between biodiversity conservation and timber production. Tree-related Microhabitats (TreMs) are used as a proxy to evaluate forest habitat value, while timber production is assessed by the number of harvested trees, biomass removal and economic income. This study takes an innovative approach by investigating TreMs using the Marteloscope in mixed Mediterranean forest. The main findings from this paper confirm that tree-related microhabitats can be considered ecological indicators effective in identifying important habitat trees, to assess forest habitat value and support tree marking for thinning operations and management.
Keywords
Biodiversity Conservation, Sustainable Forest Management, Microhabitats, Habitat Tree, Marteloscope, Timber Production, Precision Forestry
Introduction
Forests provide an array of ecosystem services of vital importance for human society ([23]). At European level, forest ecosystems cover approximately one third of the European land surface ([11]) and provide habitats and niches for a multitude of living organisms, including plants, animals, fungi, and microorganisms ([16]). European forest ecosystems are characterized by a high diversity of characteristic tree species, with 81 different habitat types identified according to the European Union (EU) Habitats Directive (92/43/ECC), and the objectives for which they are managed ([9]). They are also very diverse in terms of forest ownership and management systems ([38], [37]).
This is particularly evident in the Mediterranean basin, which is recognized as one of the most significant biodiversity hotspots in the world ([25]), yet such areas are under growing pressure due to climate change ([2]).
Since 1992, the increased interest in the protection of biodiversity has fostered the development of conservative management approaches, such as legally protected areas ([29]) or the adoption of non-intervention areas which, in some cases, has led to the abandonment of forest practices ([26]). Additionally, such cumulative effects have resulted in the depopulation of mountain and inner areas, with impacts on forest ecosystems, biodiversity conservation and loss of ecosystem services ([36], [21]). In contrast, emerging challenges of climate change adaptation have increased the demand for renewable energy, placing additional importance on forests to produce woody biomass for energy ([42]) and supporting the active management of forests. These challenges have triggered the development of new integrated and adaptive forest management strategies to counteract the effects of climate induced changes ([5], [14]) and, at the same time, to maintain and increase the provision of goods and services that forests can provide. Therefore, multifunctional management of forests, which promote forest biodiversity conservation, as well as generating revenue from forest harvesting, remains a critical task for forest managers and practitioners. Both the EU Biodiversity and Forest Strategies aim at halting biodiversity loss, increasing the provision of ecosystem services and strengthening the local economy by enhancing the competitiveness of forestry sector. Despite efforts to assess, promote and balance biodiversity conservation, timber production and provision of other ecosystems services ([42], [43]), this is still a challenge for both forest policy makers ([7]) and forest managers ([16], [20]). Excluding the formal protected areas (i.e., strictly reserves), where the protection and conservation tasks are the main forest management targets and where several restrictions on forest harvesting exist, the Natura 2000 sites play a crucial role for testing and implementing alternative and innovative forestry actions in order to evaluate their impacts on forests and the forestry sector. For these reasons, they represent an important network for evaluating trade-offs between biodiversity conservation, timber production and provision of other ecosystem services. Nevertheless, assessing the value of biodiversity of forests is a critical task, that is time consuming and often poorly implemented.
In the last decade, Tree-related Microhabitats (TreMs) have received increasing attention from forest academia ([45], [24], [18], [19], [13]) for supporting the assessment of forest biodiversity. TreMs, such as deadwood, cavities, injuries and wounds, bark structure, growth form or epiphytes represent significant niches for several living organisms, which enrich the biodiversity in forests ([45], [24]). The diversity, occurrence and abundance of TreMs is affected by forest ownership, management systems and forest structure, which influence tree species composition, tree age, morphology of trunk and crown ([44], [31], [13]).
Assessing the occurrence and abundance of TreMs on living trees can identify trees with high habitat value, so-called “Habitat Trees” ([8]), and therefore aid decision-making to pinpoint and manage highly valued trees for multifunctional objectives. Monitoring TreMs in the forest could yield useful information for tree marking operations aimed to balance timber production and conservation of biodiversity. Alternative tree selection strategies have been tested to evaluate productive use through the Marteloscope approach ([6], [39], [19]). The concept of Marteloscope was originally developed and tested in France, mainly in private forests ([6]). Usually the Marteloscope is a one ha square plot where all trees are mapped, measured and numbered in order to simulate various forestry scenarios, which are designed and implemented through a set of different tree marking selections aimed at providing a baseline to assess the resulting impacts on the forest stand. This approach is considered useful not only to evaluate human tree selection approaches but also to train foresters (and students) in order to compare the potential impacts on the provision of ecosystem services ([39], [40]). Recently, the Marteloscope approach has found an additional application for assessing the impacts of tree marking selection on the loss of TreMs ([19]).
Whether TreMs are useful ecological indicators for balancing the provision of forest ecosystem services is still being explored. However, using TreMs as an indicator of the habitat value of trees and forests could support forest managers and technicians to balance timber production with biodiversity conservation, which represents one of the most important goals of sustainable forest management.
This study aims to stress that TreMs are useful for assessing the habitat value of trees and for optimizing the provision of two contrasting forest ecosystem services, such as biodiversity conservation and timber production. This is the first study for Mediterranean mixed forest using a Marteloscope approach to investigate on TreMs. Four different silvicultural treatments have been simulated within the Marteloscope plot and the loss of TreMs has been considered for assessing the trade-off between biomass removal and habitat value of the forest stand. These simulations can give a wide range of theoretical strategies that can be implemented in practical silvicultural interventions. The study also builds on the hypothesis that indicators such as TreMs can serve as a proxy to increase biodiversity in managed forests, or at least to enhance the balance between biodiversity and timber production.
The outcome of this work should provide decision-making support to forest practitioners and forest owners in balancing management objectives, such as biodiversity conservation, timber production and forest profitability.
Materials and methods
Study area
The study area, Bosco Pennataro (Fig. 1), is located in the Molise region (Central Italy), in the Municipality of Vastogirardi (Isernia) within a Site of Community Importance “Bosco Monte di Mezzo - Monte Miglio - Pennataro - Monte Capraro - Monte Cavallerizzo”. The mean altitude of this area is 930 m a.s.l., with a mean annual temperature of 8.5 °C, and mean annual precipitation of approximately of 1012 mm. Although the forest owner is the Molise region, the forest is a State forest and was recently included in the Man and Biosphere (MaB) reserve Collemeluccio-Montedimezzo Alto Molise. The natural forest community is Aremonio agrimonioidis - Quercetum cerridis ([4]) and is classified as Oak-hornbeam forest type ([1]). The recognised historical and cultural values of Bosco Pennataro have encouraged a shift from productive to conservative forest management objectives. Nevertheless, specific silvicultural treatments are scheduled in the forest management plan to prevent pests, diseases and forest fires, as well as supporting scientific study. The current management system is high forest with continuous canopy cover and uneven aged trees including 13 different species (Fig. 2). The selected area is part of a Marteloscope demonstration site network, which was developed in the course of the Integrate+ project ([19]). To date, more than 40 Marteloscope sites have been established in 9 European countries.
Fig. 1 - The study area (represented by a one-hectare plot, see the 3D forest stand representation in the box at the bottom left) falls within the core area of the “Collemeluccio-Montedimezzo Alto Molise” Man and Biosphere (MaB) reserve and it is part of the Natura 2000 site (IT7212124)”.
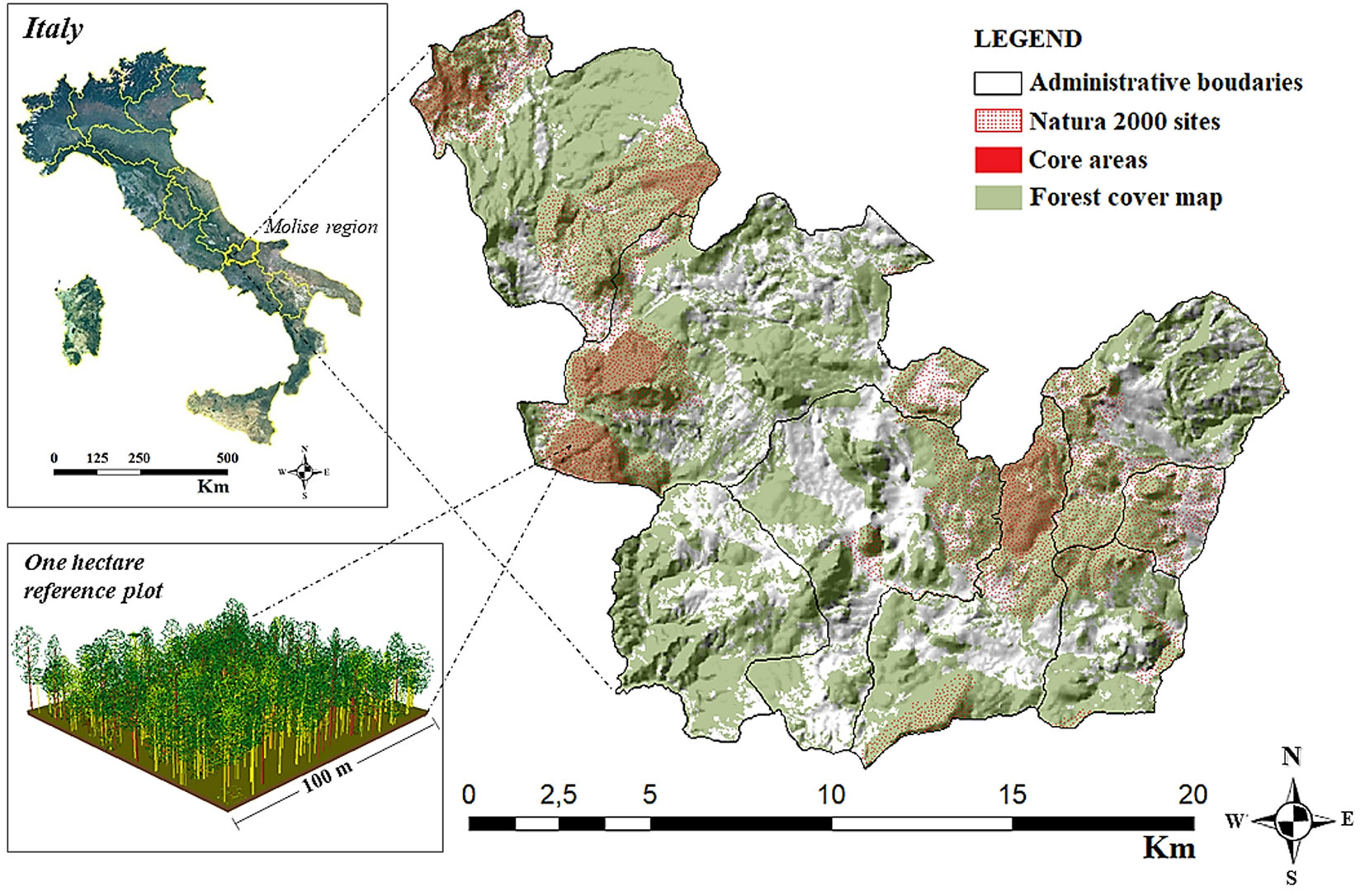
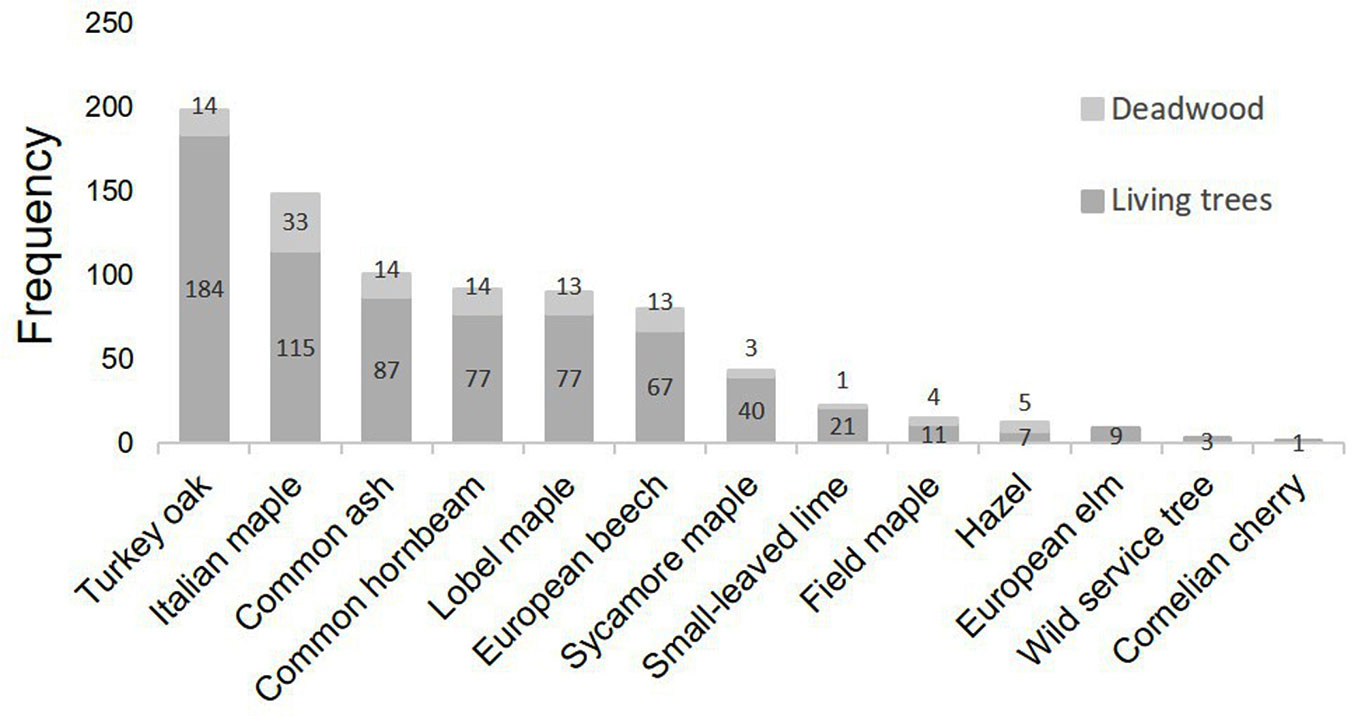
Fig. 2 - Frequency distribution of tree species within the one-hectare plot. A total of 813 trees were found in the plot, of which 699 living trees and 114 standing dead trees. For each species the number of living and dead trees is displayed in the bars.
On a regional scale, the main market for oak forests timber is firewood. According to local experiences (Alto Molise mountain district) there are mainly two firewood assortments (Tab. 1), with the average price of 37.00 EUR ton-1.
Tab. 1 - Fuelwood type and price according to the local experience for Mediterranean oak forests in Alto Molise district (Central Italy).
| Fuelwood type | Lengths (m) |
Logs diameter (cm) | Price (€ ton-1) |
|---|---|---|---|
| Harvested by chainsaw extracted by horses, processed into firewood | 1 | 3-6 | 30.00 |
| Harvested by chainsaw, skidded by horses, processed into firewood | 3 | > 6 | 44.00 |
Ground data
Ground data was collected through a field survey conducted between May and June 2016 in the one-hectare reference plot (latitude: 41° 44′ 10″ N; longitude: 14° 12′ 35″ E), namely the Marteloscope “Pennataro” (Fig. 1). All trees with diameter at breast height (DBH) > 7.5 cm were measured using Field-Map technology (⇒ http://www.fieldmap.cz/) for several attributes: stem position, species, height, diameter, crown length, crown projection and vitality.
Field-Map is an integrated tool designed for computer aided field data collection, consisting of a laser rangefinder (Forest Pro, Laser Technology, Inc., USA), with distance accuracy 3-5 cm, electronic compass (MapStar II, Laser Technology, Inc., USA), azimuth accuracy ± 0.3° and field computer equipped (Hammerhead XRT) with Field-Map Bundle ver. 13 software solution (IFER-MMS, Czech Republic). The collection of data was achieved through a Field-Map Data Collector, which records information and maps the position within the plot. The GPS Trimble GeoXT with Hurricane Antenna (sub-meter accuracy) was used to record the coordinates of the origin of the plot in order to allow a shift from local to global coordinates. The basal area (G), the volume (V) and the frequency of TreMs were calculated for each tree, as well as the species of both living and dead standing trees.
In addition, the total amount of growing stock was calculated according to Federici et al. ([10]).
TreMs inventory
TreMs are usually referred to as tree structures ([45]), which develop under specific conditions or following particular disturbance events (e.g., storm, fire). Their development can also be strongly influenced by silvicultural interventions and forest management aims ([13]). The assessment of TreMs in the plot was carried out using a tree microhabitat reference field-list ([15]). The original 63 TreMs of the field-list are grouped into 7 categories (Tab. 2) in order to present the results and the discussion in an accessible format.
Tab. 2 - List of the tree-related microhabitats (TreMs) and their ecological role.
| TreMs | Short description | Ecological role |
|---|---|---|
| Cavities | Depending on their origin, they can be distinguished in woodpecker cavities or rot holes and dendrotelms, which often accumulate water and microsoil. |
Many species use cavities for nesting, resting or wintering ([33]). Old cavities can be reused by secondary cavity users, both vertebrates and invertebrates and also epiphytic lichens and bryophytes ([12]). |
| Injuries and wounds |
Tree cleft, lightning or fire scars, broken trunk or crown, splintered stem which expose heartwood and bark loss which expose sapwood. |
Microhabitats resulting from natural wounds but also from harvesting operations. In relation to the decay type, a tree will provide characteristic tree structures over time useful for nesting, foraging in the bark and in the interior after carpenter ants ([8]). |
| Bark | Peeling off of trunk bark providing either a shelter or a pocket that can be filled with organic material; mainly occurring when trees are weakened by stress lose the bark over desiccating wood. | They are particular important for bats, birds, insects and molluscs as roosting nesting and foraging sites ([15], [45]). |
| Deadwood | This habitat occurs on living trees in the form of dead limbs in contact with living wood (xylem and phloem flow). It is a significant part of total deadwood in forest ecosystems |
Deadwood is one of the most important biodiversity indicators ([41]). Crown deadwood can be considered a key attribute for saproxylic beetle diversity ([28]). |
| Deformation and growth form | Witches’ brooms, outgrowths and epicormics shoots on the trunk or branches of trees caused by a parasite or from epicormics buds. Natural cavity at the base of the tree trunk formed by the tree roots. |
Salamanders use holes near buttress roots at the basis of trees as shelters ([3]). Cankers and burrs evolve in deadwood used by various species for many purposes ([8]). |
| Epiphytes | Fruiting bodies of fungi, moss, lichens and lianas. The epiphytic plants and conks of fungi are important ecological niches for several species. |
They are important environmental indicators ([27]). They are also indicators of silvicultural interventions ([34]). |
| Other | Include nests of invertebrates, small vertebrates and large vertebrates, as well as the result of micro-pedogenesis from epiphytic mosses, lichens or algae and fresh significant flow of sap and resin. |
Trees are used for the construction of nests of large birds ([30]). Many xylophilous insects are specialized in fresh flow of sap and resin (see [13]). |
During the field survey, all TreMs were recorded individually on every single living tree and on standing dead trees, stumps and snags included.
Intervention scenarios
Four different harvesting interventions were simulated to compare the trade-offs between timber production, net revenue and biodiversity conservation. We hypothesized the adoption of the selection system based on management objectives, which require the retention of some large trees on-site for aesthetics and/or as wildlife habitats while allowing increased volume gains for future timber harvesting. The proposed silvicultural interventions aim to decrease the current basal area value by 30% at stand level, reducing competition and reinvigorating tree growth. The criteria used for defining the scenarios were: (i) crown damages/vitality; (ii) occurrence of TreMs; (iii) tree height; (iv) tree DBH; and (v) slenderness coefficient. The combination of these criteria enabled the simulation of tree marking operations (Tab. 3). Crown damage is combined with tree height, tree DBH and slenderness coefficient, respectively in scenarios 1, 2 and 3 (henceforth: S1, S2, S3). These scenarios are production oriented, while S4 combines trees lacking TreMs with tree DBH, thus representing a conservation-oriented scenario.
Tab. 3 - Forestry scenarios (harvested trees) simulated in this work.
| Scenario | Criterion 1 | Criterion 2 |
|---|---|---|
| S1 | All the trees with crown damages | Tree height - From highest to lowest |
| S2 | All the trees with crown damages | Tree DBH - From largest to smallest |
| S3 | All the trees with crown damages | Trees slenderness coefficient - From highest to lowest |
| S4 | All the trees without TreMs | Tree DBH - from largest to smallest |
The trade-off assessment was carried out using the following criteria: (i) timber production; (ii) number of harvested trees; (iii) economic income; (iv) number of TreMs removed. Timber production refers to the amount of harvested growing stock available from the four scenarios. Number of harvested trees reflects the quantity of trees that need to be harvested per scenario to reduce the basal area by 30%. Economic income represents the profitable value (in EUR) derived from marketable firewood. Finally, number of TreMs removed refers to the reduction of TreMs due to harvesting operations and thus the effects on biodiversity within the forest stand.
Results
Plot inventory
The survey recorded a total of 813 standing trees over 13 different species (Fig. 2) of which 699 are living and 114 are standing dead trees. Living trees also include trees with some damage in the canopy (e.g., dead branches) for less than 1/3 of the crown (34 trees), between 1/3 and 2/3 of the crown (14 trees) and more than 2/3 of the crown (7 trees). Standing dead trees include 49 snags and 14 stumps. The uneven-aged stand has heterogeneous but continuous canopy cover and continuous vertical tree distribution ([22]). The study area is characterised by high variability, both in terms of number of tree species and DBH distributions (Fig. 3).
Fig. 3 - Boxplot showing the main statistics (minimum, first quartile, median, third quartile, and maximum) of tree DBH distribution for all living trees in the study area. Maples were grouped in a single category, while “Other” indicates the remaining 4 tree species. Circles and stars are outliers and extreme outliers, respectively. (n): number of trees for each species or group of species. Most of oak and beech trees (~75% and ~50%, respectively) had a DBH larger than the majority (~75%) of the remaining species.
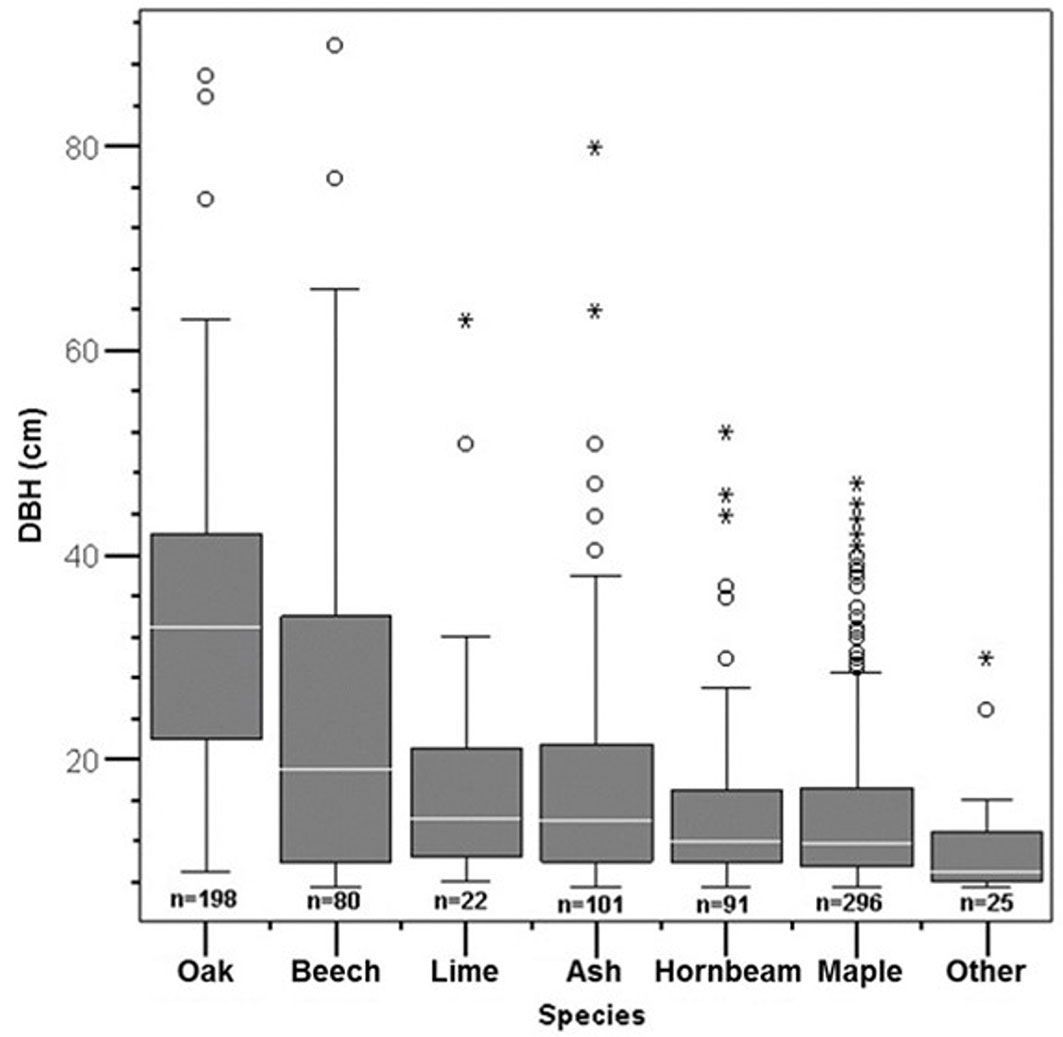
Turkey oak (Quercus cerris L.) is the primary tree species as for abundance (Fig. 2, Fig. 3), basal area and volume (Tab. 4). Beech (Fagus sylvatica L.) is the secondary species in terms of volume and mean DBH, though its abundance is lower than that of maples (Acer spp., including several species), ash (Fraxinus excelsior L.) and hornbeam (Carpinus betulus L.). Results show that the main canopy layer is dominated by oak and beech, due to their large DBH (Fig. 3), while the remaining species occupy secondary canopy layers.
Tab. 4 - Main stand structural characteristics in the study area. Detailed data for all the Marteloscope plots are available in the Kraus et al. ([17]) database.
| Species | Living trees | Standing dead trees | |||||||
|---|---|---|---|---|---|---|---|---|---|
| n | DBH (cm) | V (m3 ha-1) |
G (m2 ha-1) |
TreMs (n) | n | DBH (cm) | V (m3 ha-1) |
TreMs (n) | |
| Maple | 243 | 16 | 51.2 | 6.3 | 197 | 53 | 12 | 3.6 | 50.0 |
| Hornbeam | 77 | 16 | 19.5 | 2.0 | 35 | 14 | 11 | 0.6 | 5.0 |
| Beech | 67 | 24 | 45.6 | 4.4 | 58 | 13 | 31 | 9.3 | 19.0 |
| Ash | 87 | 19 | 33.8 | 3.6 | 73 | 14 | 15 | 0.9 | 6.0 |
| Other | 20 | 12 | 1.9 | 0.3 | 2 | 5 | 8 | 0.1 | 1.0 |
| Oak | 184 | 34 | 205.2 | 18.8 | 283 | 14 | 20 | 4.5 | 14.0 |
| Lime | 21 | 19 | 8.0 | 0.9 | 11 | 1 | 9 | 0.0 | 0.0 |
| Total | 699 | 20 | 365.3 | 36.3 | 659 | 114 | 15 | 19.0 | 95 |
The growing stock is 384.3 m3 ha-1 of which 365.3 m3 ha-1 are living trees and 19.0 m3 ha-1 are standing dead trees (Tab. 4). In contrast with the living tree abundance, beech is the species with the highest volume of standing dead trees (9.3 m3 ha-1) followed by oak (4.5 m3 ha-1). Results also show that maple is the species with the largest number of standing dead trees, despite inferior DBH, with a cumulative volume of 3.6 m3 ha-1.
TreMs occurrence
A total of 754 TreMs were recorded in both living trees and standing deadwood (Tab. 5). Among living trees, oak trees had the highest number of TreMs. Despite a smaller DBH, maple trees showed the highest number of TreMs among the standing dead trees. Epiphytes (e.g., lianas, epiphytic bryophytes, foliose and fruticose lichens) and cavities (e.g., branch holes, dendrotelms and water-filled holes) are the most abundant TreMs recorded in this study, 25% and 41% respectively. Dead crown, such as dead branches and limbs, is also a recurrent microhabitat (10%), particularly in oak trees.
Tab. 5 - Abundance (n ha-1) of TreMs by species and microhabitat types. The 63 TreMs were grouped in 6 main categories. The overall average of TreMs per trees is 0.9, with standard deviation of 1.42 and standard error of 0.05. The maximum value of TreMs per tree was 11, while for 441 trees no microhabitats were detected.
| Species | Cavities | Injuries | Deadwood | Bark | Growth form |
Epiphytes | Others | TreMs |
|---|---|---|---|---|---|---|---|---|
| Maple | 93 | 7 | 19 | 15 | 19 | 76 | 18 | 247 |
| Hornbeam | 20 | 2 | 3 | 1 | 5 | 5 | 4 | 40 |
| Beech | 45 | 4 | 8 | 2 | 4 | 7 | 7 | 77 |
| Ash | 32 | 4 | 8 | 2 | 4 | 22 | 7 | 79 |
| Other | 1 | 0 | 0 | 1 | 0 | 1 | 0 | 3 |
| Oak | 111 | 9 | 42 | 1 | 25 | 78 | 31 | 297 |
| Lime | 4 | 0 | 1 | 1 | 2 | 2 | 1 | 11 |
| Total | 306 | 26 | 81 | 23 | 59 | 191 | 68 | 754 |
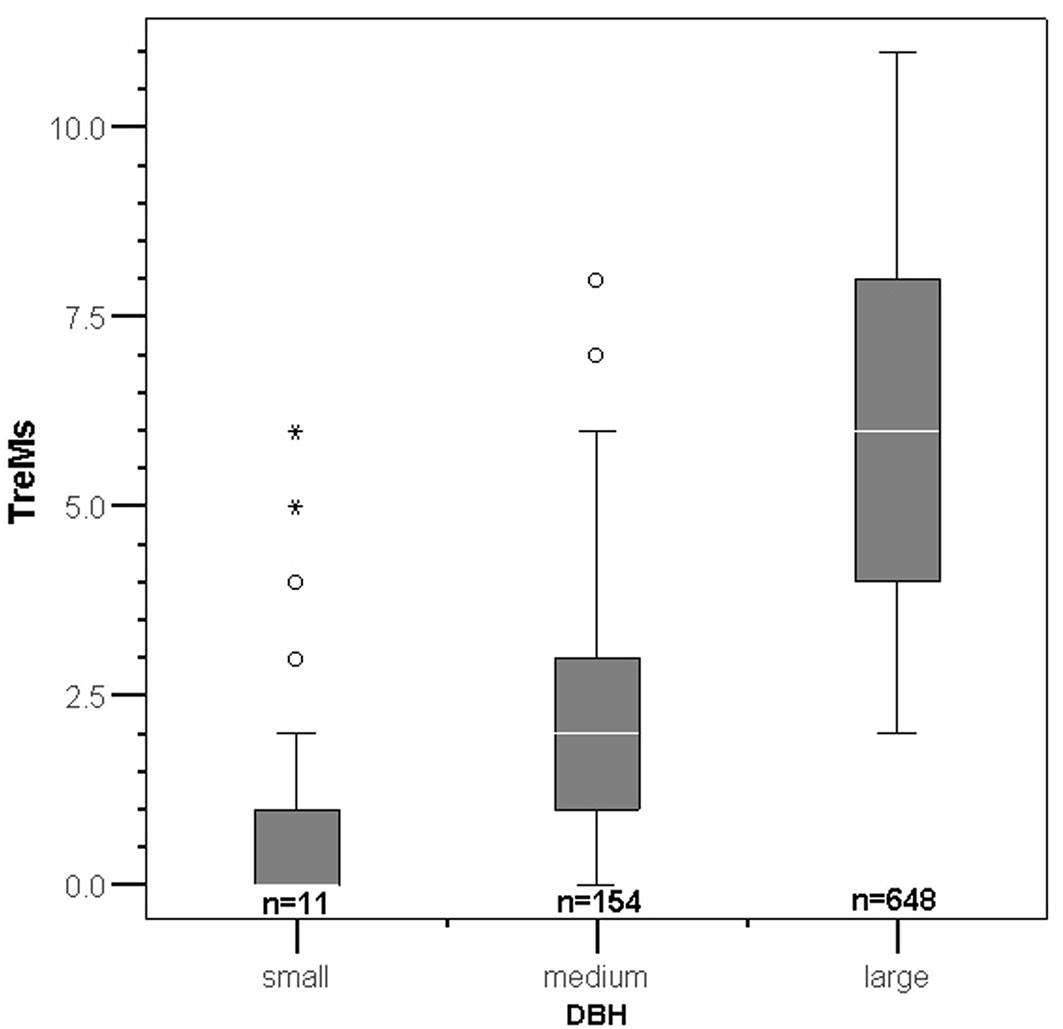
TreMs occurrence showed an increasing trend with DBH (Fig. 4); with the exception of some outliers and extreme outliers, trees with small DBH present no more than 2 TreMs on an individual tree.
Fig. 4 - TreMs frequency by DBH classes. (small): tree DBH <30cm; (medium): 30cm < DBH < 60 cm; (large): DBH >60 cm. The white line in the middle of each box is the median of TreMs abundance per individual tree. The top and the bottom of boxes indicate the 75th and 25th percentile, respectively. Circles and stars are outliers and extreme outliers, respectively.
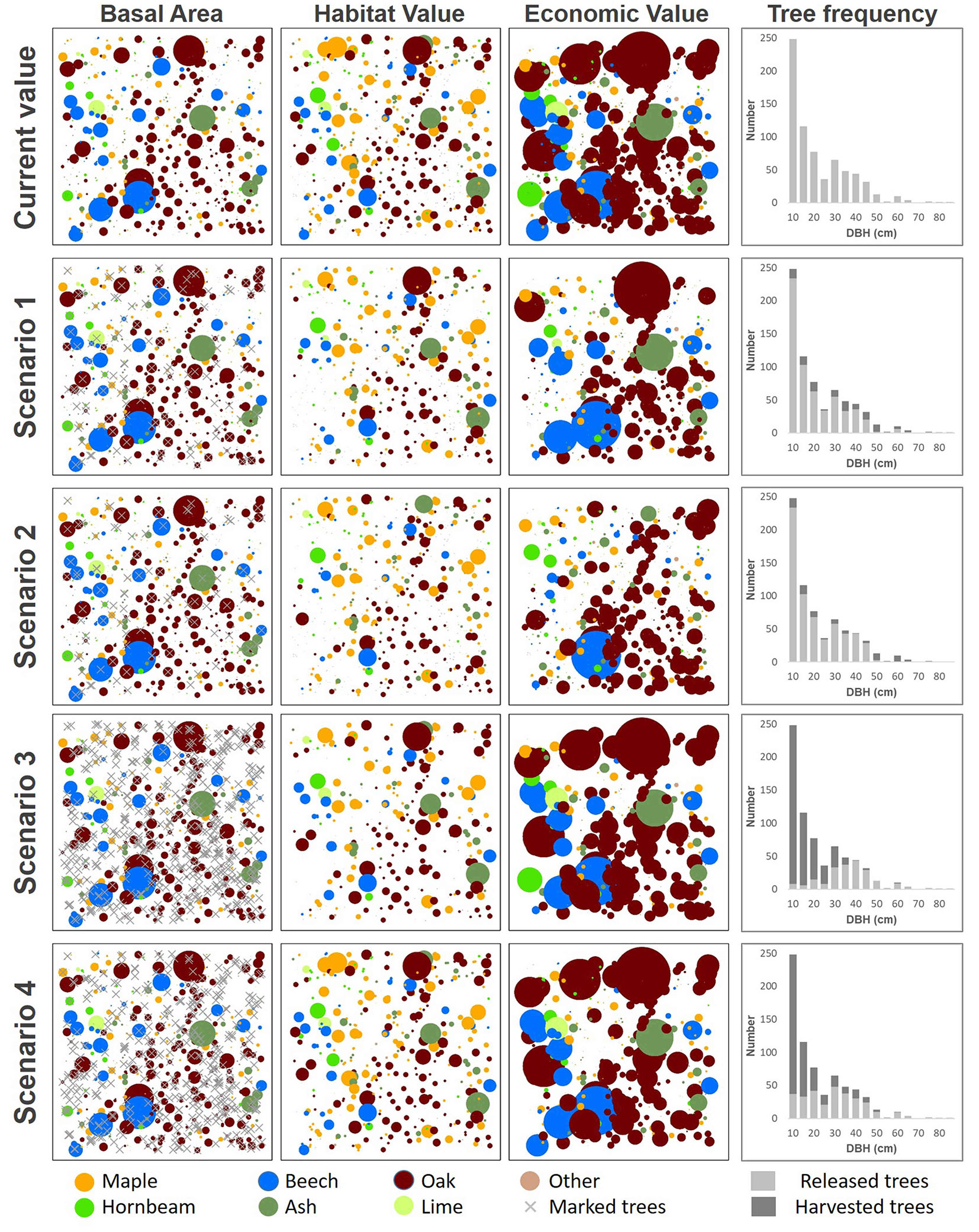
The habitat value of trees within the study area (Fig. 5) reflects the abundance and spatial distibution of TreMs on habitat trees. Although large trees present more TreMs than small trees in absolute terms, differences between economic and habitat values are highlighted for small trees, especially maples.
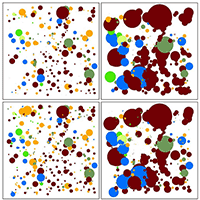
Fig. 5 - Forestry intervention scenarios showing the spatial distribution of basal area, habitat value and economic value. The colour reflects the species (see the bottom legend), while the dimension of the circle reflects the value of: (1st column) basal area (m2 ha-1); (2nd column) habitat value (abundance of TreMS per individual tree); (3rd column) Economic Value (EUR per tonnes). The histograms in the 4th column show the tree frequency by DBH classes (interval 5 cm) according the current situation and after the four simulated interventions.
Forestry scenarios
In the study area (1 ha), basal area of living trees was 36.3 m2 and total aboveground biomass was 332.3 tons. The economic value corresponded to about 11.210 euros for all living trees. Over the four interventions scenarios, harvesting impacted the forest structure, abundance and spatial distribution of TreMs, as well as the economic and habitat values of the forest stand (Fig. 5). It should be noted that selecting trees for harvesting based on DBH has a larger impact on the loss of TreMs than using height as thinning criterion, as demonstrated by S2. Regarding forest structure, the four simulated harvesting scenarios involved an obvious change in the frequency distribution of tree DBH (Fig. 5), particularly in S3 and S4, where it significantly differed from the original distribution, reflecting the shift from the current uneven-aged to an even-aged structure.
Fig. 6 highlights the various impacts of the different interventions by comparing harvesting intensity, loss of TreMs, above ground biomass and economic revenue. S1 and S2 are the most suitable scenarios in regard to the effective harvesting, as they provide a high financial return, although a reduced number of trees (<200 out of 659 in both cases) were removed. Contrastingly, in S3 a large number of trees with low diameters is removed, resulting in the lowest economic value. Finally, trees hosting microhabitats are not harvested in the scenario S4; however, a relatively high number of trees is removed, resulting in a considerable economic return, though the selective nature of harvesting operations increases the cost of intervention.
Fig. 6 - Trade-offs comparison among the simulated scenarios (S1, S2, S3 and S4). No. of harvested trees, loss of TreMs and tons of timber are displayed for each simulated intervention. The black line describes the economic value in EUR of firewood obtainable from the different scenarios. S4 showed the highest contribution to biodiversity conservation.
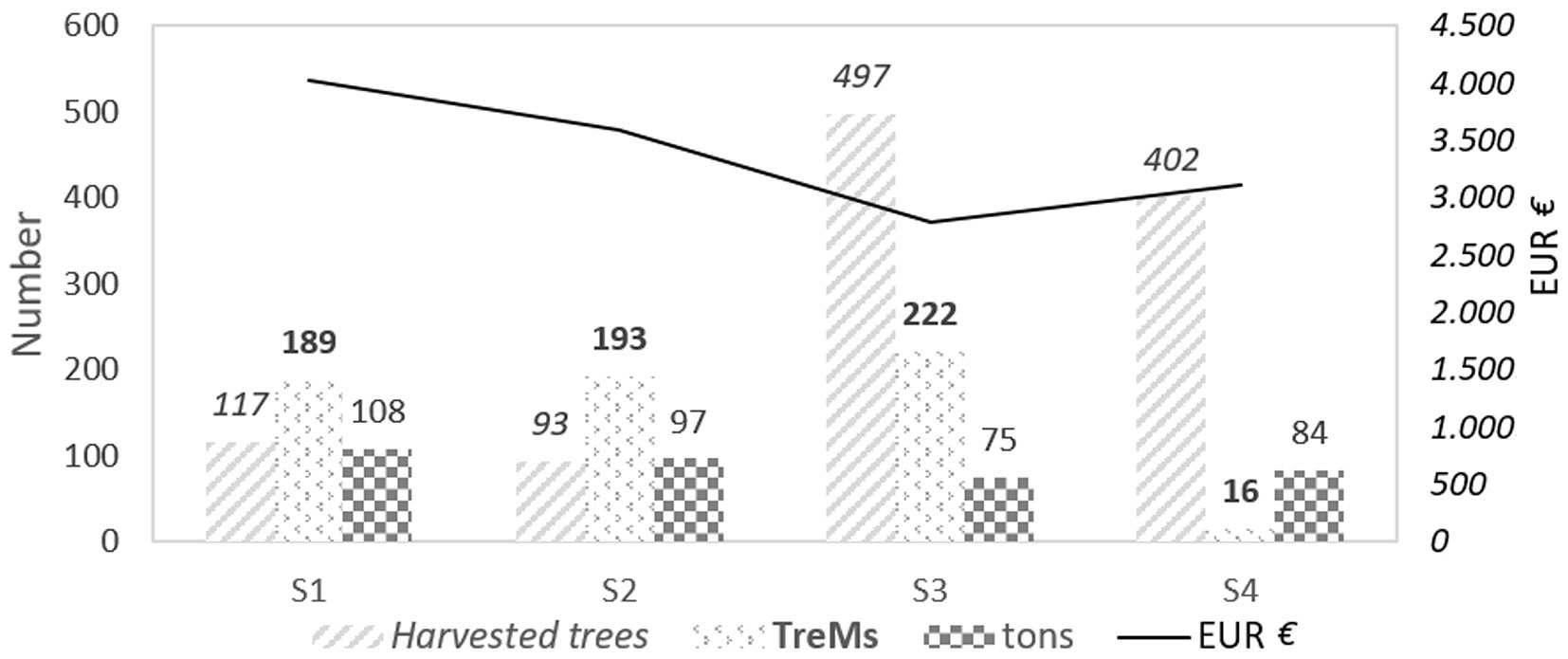
S4 is the most suitable intervention for ecological objectives, retaining the largest number of TreMs (only 16 were removed). In contrast, scenarios S3, S2 and S1 resulted in the removal of 222, 193 and 189 TreMs, respectively. Therefore, the scenario S4 represents a viable management option, especially considering that the experimental plot is located in the MaB reserve core area and is designated as a Natura 2000 site.
Discussion
Habitat value and economic return were the key trade-off evaluations of this work. The habitat value was evaluated by the occurrence and abundance of microhabitats on tree stems, branches and crowns, whereas the economic value was evaluated by considering both the harvesting effort and the value of firewood according to the local wood market. We found an increase in the occurrence and abundance of TreMs with the increment of tree diameter, as already reported by previous studies ([44], [31]). As large trees are normally associated with a higher frequency of TreMs, we can state that the TreMs assessment could represent a viable method to identify and quantify potential conflicts in forest management goals. Indeed, the retention of TreMs hinders the maximisation of the economic revenue during harvesting operations, as demonstrated in this study. In order to counterbalance these contrasting aims, the identification of TreMs will help forest managers to develop more informed decisions during tree marking operations. Nevertheless, further research on the relationship between TreMs occurrence, forest structure and silvicultural interventions is needed to assess whether or not the occurrence of TreMS depends on the traits of tree species or on silvicultural choices. For example, growth forms often depend on specific tree species and age ([24]), while damage caused by harvesting operations or storms could increase the occurrence of tree injuries, deadwood and bark loss. In our study area, cavities were the most abundant TreMs, while bark loss and injuries were least frequent. The low frequency of bark loss and injuries is due to the scarcity of management interventions in the study area, closely related to forest fire prevention and scientific purposes (i.e., Life + ManFor C.BD.). The high occurrence of epiphytes also confirms a low level of disturbance in the study forest ([34]). Normally, a higher number of TreMs are detected in unmanaged rather than in managed stands ([13]). Nevertheless, active forest management can also increase TreMs abundance by safeguarding habitat trees from harvesting operations. In addition, active forest management can influence the spatial distribution of TreMs ([44]) by identifying and removing potential competitors that would hinder timber production and/or TreMs occurrence. This is particularly important for timber production forests, where the designation of habitat trees within the stand (due to their high habitat value) should involve their exclusion from harvesting, thereby balancing the increasing demand for wood energy with biodiversity conservation objectives. Thus, TreMs can be considered helpful ecological indicators for assessing the habitat value of forests and supporting managers and practitioners in their tree marking operations.
The four scenarios simulated in this study show four different outcomes obtained by balancing economic benefits with ecological constraints. The scenario S1 was the most profitable, yielding the highest economic benefits while retaining 71.3 % of the original number of TreMs. S2 simulated the lowest cutting intensity among the four kind of interventions, and was the most balanced scenario, achieving the second most profitable revenue while preserving 70.7 % of the original number of TreMs. In this case, the cutting effectiveness reduces the harvesting costs and ensures a considerable removal of above ground biomass and an expected high financial return. The scenario S3 was the least beneficial in terms of economic return, as the removal of a large number of trees with greater slenderness coefficient increases the costs of harvesting operations. More importantly, the high cutting intensity affects the habitat value of the forest and strongly influences the stand structure that shifts from uneven-aged to even-aged. Furthermore, this scenario had the low biodiversity retention in term of TreMs (33.7 %) among the four intervention types. Finally, the scenario S4 proved ecologically effective as trees with high habitat value (i.e., TreMs) are excluded from harvesting. This scenario also provides a relatively significant economic profit, as well as an enhancemet of the habitat value. Based on our results, S4 is an appropriate level of intervention for forests in protected areas (e.g., Natura 2000 forests) where the main aim is biodiversity conservation or in particular forests where scenic beauty is more important than timber production. TreMs could thus be considered as helpful ecological indicators when applying close-to-nature-forest systems.
In light of the above results, we can conclude that biodiversity conservation and timber production objectives in forest management can be reconciled. However, expertise and local knowledge of forest practitioners is required, as well as a strengthened cooperation between forest academia, practitioners and managers.
A more comprehensive analysis of similar interventions (S1-S4) throughout the Integrate+ Marteloscope demonstration site network ([19]) will help our understanding of the range of TreMs among different forest types and forest management systems across Europe. This paper contributes 3 key observations:
- the wide range of microhabitats calls for a better evaluation of trees during tree marking operations in order to support the biodiversity conservation within and outside Natura 2000 forest sites;
- habitat trees can be easily integrated into forest management and support the provision of ecological niches for many endangered species, linear green infrastructures, as well as urban forests, which are rarely considered under the Natura 2000 directives and sustainable forest management indicators;
- forest management can actively enhance the conservation of biodiversity in forests, evidencing and valuing the multifunctional role of forests.
As training sites Martlescopes can provide a significant contribution in familiarising forest practitioners and managers with balanced decision-making (e.g., recognising tree characteristics, and a range of simulated silvicultural interventions). Continuous improvements in the detection of indicators for sustainable forest management focused on the use of remote sensing techniques and precision forestry should be further supported by monitoring and management activities.
A recent study from the Swiss Federal Research Institute WSL shows how data from Terrestrial Laser Scanning could be useful for identifying TreMs, through their inclusion in the National forest inventory ([32]). Additionally, recent improvements in the use of Airborne Laser Scanning data for monitoring forest resources, especially single tree detection, could be useful for mapping tallest and largest trees that often correspond to habitat trees. Although further improvements and testing are necessary, promising results have been reported for uneven-aged forests, where the stratification of canopy allowed for the discrimination of trees at the emergent canopy layer, including trees at the middle and under canopy layers ([35]).
Conclusions
Despite the relatively small study area, our study demonstrates that TreMs can be used as ecological indicators to identify habitat trees, which can subsequently be used to assess forest habitat value and enhance tree marking operations. Nevertheless, further work is needed to: (i) assess the spatial distribution of TreMs within forest stands; (ii) increase the evaluation accuracy of habitat value; (iii) identify TreMs hotspots; and (iv) apply spatial attributes (e.g., tree group selection) to simulate forestry scenarios rather than only using dendrometric variables.
This study demonstrates that TreMs are viable indicators which can be used to reconcile habitat and economic values, thus providing a useful insight for decision-makers, forest practitioners and managers in order to implement a sustainable forest management.
Acknowledgments
We would like to thank the Dr. Euan Bowditch from Inverness College UHI (Scotland, UK) and Nina Sassani from Roxburgh College (Victoria, Australia) for their great contribution in proofreading and supporting our work and improving the quality of this paper.
This research was supported by the LIFE program and the Italian Academy of Forest Sciences, in the framework of the project “FRESh LIFE - Demonstrating Remote Sensing integration in sustainable forest management” (LIFE14 ENV/IT/000414). Support has also been provided by the German Federal Ministry of Food and Agriculture (BMEL), through the “Integrate+” project which established a European network of demonstration sites for the integration of biodiversity conservation into forest management.
References
Online | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Marco Marchetti
Centro di Ricerca per le Aree Interne e gli Appennini (ArIA), Università degli Studi del Molise, v. de Sanctis, I-86100 Campobasso (Italy)
Bruno Lasserre
Marco Marchetti
Dipartimento di Bioscienze e Territorio, Università degli Studi del Molise, c.da Fonte Lappone, I-86090 Pesche, IS (Italy)
Chair of Silviculture, University of Freiburg, Tennenbacherstr. 4, D-79085 Freiburg im Breisgau (Germany)
European Forest Institute, Platz der Vereinten Nationen 7, D-53133 Bonn (Germany)
Corresponding author
Paper Info
Citation
Santopuoli G, di Cristofaro M, Kraus D, Schuck A, Lasserre B, Marchetti M (2019). Biodiversity conservation and wood production in a Natura 2000 Mediterranean forest. A trade-off evaluation focused on the occurrence of microhabitats. iForest 12: 76-84. - doi: 10.3832/ifor2617-011
Academic Editor
Marco Borghetti
Paper history
Received: Aug 29, 2017
Accepted: Nov 04, 2018
First online: Jan 24, 2019
Publication Date: Feb 28, 2019
Publication Time: 2.70 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2019
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 51680
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 39197
Abstract Page Views: 6331
PDF Downloads: 5122
Citation/Reference Downloads: 22
XML Downloads: 1008
Web Metrics
Days since publication: 2724
Overall contacts: 51680
Avg. contacts per week: 132.80
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2019): 27
Average cites per year: 3.86
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Review Papers
The forest biodiversity artery: towards forest management for saproxylic conservation
vol. 9, pp. 205-216 (online: 26 October 2015)
Research Articles
Early responses of biodiversity indicators to various thinning treatments in mountain beech forests
vol. 11, pp. 609-618 (online: 25 September 2018)
Research Articles
Factors affecting the quantity and type of tree-related microhabitats in Mediterranean mountain forests of high nature value
vol. 14, pp. 250-259 (online: 21 May 2021)
Research Articles
The effects of forest management on biodiversity in the Czech Republic: an overview of biologists’ opinions
vol. 15, pp. 187-196 (online: 19 May 2022)
Book Reviews
National forest inventories: contributions to forest biodiversity assessments (2010)
vol. 4, pp. 250-251 (online: 05 November 2011)
Research Articles
Investigating the effect of selective logging on tree biodiversity and structure of the tropical forests of Papua New Guinea
vol. 9, pp. 475-482 (online: 25 January 2016)
Review Papers
Biodiversity assessment in forests - from genetic diversity to landscape diversity
vol. 2, pp. 1-3 (online: 21 January 2009)
Research Articles
Bird response to forest structure and composition and implications for sustainable mountain forest management
vol. 19, pp. 18-27 (online: 11 January 2026)
Research Articles
Rewilding beech-dominated temperate forest ecosystems: effects on carbon stocks and biodiversity indicators
vol. 18, pp. 1-9 (online: 02 February 2025)
Short Communications
An approach to measuring biodiversity and its use in analysing the effect of nitrogen deposition on woodland butterfly populations in the Netherlands
vol. 2, pp. 46-48 (online: 21 January 2009)
iForest Database Search
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords







