
Contrasting multi-taxa diversity patterns between abandoned and non-intensively managed forests in the southern Dolomites
iForest - Biogeosciences and Forestry, Volume 10, Issue 5, Pages 845-850 (2017)
doi: https://doi.org/10.3832/ifor2181-010
Published: Oct 26, 2017 - Copyright © 2017 SISEF
Research Articles
Abstract
The abandonment of silvicultural activities can lead to changes in species richness and composition of biological communities, when compared to those found in managed forests. The aim of this study was to compare the multi-taxonomical diversity of two mature silver fir-beech-spruce forests in the southern Dolomites (Italy), corresponding to the European Union habitat type 9130. The two sites share similar ecological and structural characteristics, but differ in their recent management histories. In the last 50 years, one site underwent non-intensive management, while the other was left unmanaged and was included in a forest reserve. The species richness and composition of eight taxa were surveyed in the two sites between 2009 and 2011. The difference in mean species richness between the two forest management types was tested through permutation tests, while differences in species composition were tested by principal coordinates analysis and the permutational multivariate analysis of variance. Mean species richness of soil macrofungi, deadwood lichens, bark beetles, and longhorn beetles were significantly higher in the abandoned than in the non-intensively managed forests. Deadwood fungi and epiphytic lichens did not differ in mean species richness between the two study sites, while mean species richness of ground beetles and birds were higher in the non-intensively managed than in the abandoned forest. Significant differences in species composition between the two sites were found for all the taxa, except for longhorn beetles. These results indicate that improving forest landscape heterogeneity through the creation of a mosaic of abandoned and extensively managed forests should better fulfill the requirements of ecologically different taxa.
Keywords
Asperulo-Fagetum, Forestry Abandonment, Biodiversity Conservation, Selection Cutting, Natura 2000, Silver Fir
Introduction
Intensive forest management practices are frequently a major driver of biodiversity loss worldwide. In Europe, a large body of research has been conducted to evaluate the impact of forestry on biodiversity in European forests, and provide science-based guidelines for conservation. Many of these studies are related to boreal forests ([32]), and are based on comparisons between intensively managed forests (e.g., those subjected to clearcutting) and forests managed for conservation (e.g., old-growth forests, minimal intervention forests, or stands where forestry practices have ceased in the last decades - [18]). From these studies, there is general agreement regarding the beneficial effects of forest abandonment on biodiversity ([32]), due to the development of habitat attributes which are more similar to those of old-growth forests ([51]).
However, the positive effects of forest abandonment may not be consistent across taxa, especially when the transitions of non-intensively managed forests are considered. According to the intermediate disturbance hypothesis ([19], [9]), higher species diversity may be expected under non-intensive forest management systems where disturbances occur at intermediate frequencies (e.g., selection systems), than would be expected in an entirely unmanaged forest ([48]).
This framework could apply to several forests in the European Alps, which have been non-intensively managed for both timber production and multiple ecosystem services since the second half of the 20th century ([35]). For example, in Alpine larch (Larix decidua Mill.) woodlands, forest abandonment could be detrimental for species which benefit from intermediate disturbance regimes ([26]). However, despite the fact that alpine forests are important conservation habitats in Europe, with many included in the Natura 2000 network, multiple taxa biodiversity effects of forest abandonment as compared to non-intensive forest management are not well understood.
Assessing the response of multiple taxa to forest management could provide more robust information on forest biodiversity patterns and drivers compared to focusing only on a single taxon ([21], [52], [13]). For example, the use of a multi-taxon approach is the basis for elucidating the potential co-variation among organisms exploiting different substrates (e.g., soil, dead wood, tree bark) and occupying different trophic levels ([6]). Information on species diversity patterns for multiple taxa is therefore crucial for the assessment of conservation priorities ([33]) and of surrogate taxa for the representation of biodiversity ([23], [40], [17]). The need for these data is currently one of the key challenges in forestry research ([6], [15]).
Currently, the knowledge gap regarding the effects of forest management on multiple taxa is related to two main issues: (a) the complexity in controlling potential confounding factors, such as site conditions, landscape context, past management history ([42]), that hinders the development of studies with adequate replication for general statistical inference ([33]); and (b) the cost of performing multi-taxon studies that simultaneously evaluate the response of different taxa which have a range of ecological requirements ([32]).
In this study, we tested the hypothesis that the effects of forest abandonment on biodiversity are consistent and positive across taxa in non-intensively managed forests. We surveyed eight taxa in mountain forests dominated by silver fir (Abies alba Mill.), beech (Fagus sylvatica L.), and Norway spruce (Picea abies [L.] H. Karst.). We compared the diversity patterns between two neighboring watersheds in the Italian Alps, one of which has been subjected to forest abandonment for 50 years, and the second which has been non-intensively managed by selective logging. To mitigate the impact of pseudo-replication in our experimental design, the forests of the two watersheds were characterized by matched phytosociological, past management history, climatic, and topographic conditions ([42]). The eight taxa were selected to cover a wide range of resource uses and substrates, in terms of soils, deadwood and epiphytic organisms, photosynthetic organisms, decomposers, and predators.
Methods
Study areas
The study was conducted in a mountain forest district of the southern Dolomites, in the eastern Italian Alps. The mean annual temperature is 7.2 °C, and the mean annual precipitation ranges from 1300 to 1500 mm, mainly concentrated in May-June and October-November. The most common forest soils are cambisols ([4]), and the bedrock consists of calcareous and dolomitic materials ([37]).
The focus of this research was in the adjacent watersheds of Tovanella (46° 18′ N, 12° 18′ E) and Cajada (46° 14′ N, 12° 14′ E). Both watersheds are approximately 1040 ha in size, with elevations ranging from 550 to 2500 m a.s.l., and have a similar history of forest management, but are currently subjected to contrasting forest management regimes ([42]). Both forests have been heavily logged in the last century. From 1943 to 1953, the ratio of yield to increment was up to 150%. This resulted in a very low growing stock of less than 200 m3 ha-1, with a low density of large trees ([46], [22]).
In the Tovanella watershed, both forestry and grazing ceased in 1957 ([46]), and the site has been part of a nature reserve since 1971. In Cajada watershed, non-intensive logging is conducted by group-selection, with coupe sizes not larger than 1000 m2, and grazing is not admitted inside forests. Based on the data available for a forest compartment adjacent to the surveyed plots, the mean annual harvest in the Cajada watershed in the period 1991-2003 (prior to the survey time) has been 1.7 m3 ha-1, which corresponds to 0.6% of the growing stock and 33% of the annual increment ([3]). Due to these different management regimes, the forests of the two watersheds differ in some structural parameters, which are summarized in Tab. 1. Both watersheds are currently included in Natura 2000 sites (Tovanella: site IT3230031, 8845 ha, and IT3230089, 70.397 ha; Cajada: site IT3230083, 31.383 ha), and are therefore aimed at biodiversity conservation according to the Habitats Directive (92/43/EEC).
Tab. 1 - Mean (± standard deviation) structural parameters showing significant differences (P < 0.01) between sample sites at the non-intensively managed and the abandoned forests. P-values were obtained using a linear mixed model (forest management as a fixed factor, plot as a random factor) with a restricted maximum likelihood (REML) approach.
| Variable | Type | Non-intensively managed | Abandoned |
|---|---|---|---|
| Beech crown cover (%) | - | 28.6 ± 19.7 | 55.6 ± 13.8 |
| Coarse woody debris (m3 ha-1 ) |
Stumps | 23.0 ± 9.9 | 6.0 ± 3.5 |
| Snags | 1.7 ± 3.4 | 14.1 ± 8.5 | |
| Logs | 0.2 ± 0.2 | 18.5 ± 10.2 |
Sampling design and species data collection
In each watershed, ten sampling units (Fig. 1) were randomly selected and permanently marked in the field. Each sample was located within a mature mixed silver fir-beech-Norway spruce forest (EU habitat type 9130) with a slope of less than 26° ([20], [11]). At each sample location, a 12.5 m radius circular plot (491 m2) was established, with a minimum distance of 200 m between each plot. The average altitude of the plots in Cajada was 1221 ± 103 m a.s.l., with aspects ranging from NNE to S. The average altitude of plots in Tovanella was 1228 ± 43 m a.s.l, all with N aspect. Previous analyses of the vegetation units based on regional phytosociological classification confirmed that the forest stands of the two watersheds represent the same forest type ([42]). This equivalence is fundamental, given that different forest types may host different communities with different species richness and composition.
Fig. 1 - Schematic diagram of the sampling plots, where the silhouettes of animals represent the eight taxonomic groups surveyed. Beetles were surveyed with two pitfall and one window trap, while the other taxa were surveyed by counting the species occurring inside the sampling plot, on the ground, living trees, deadwood, or acoustic detection (images with different scales).
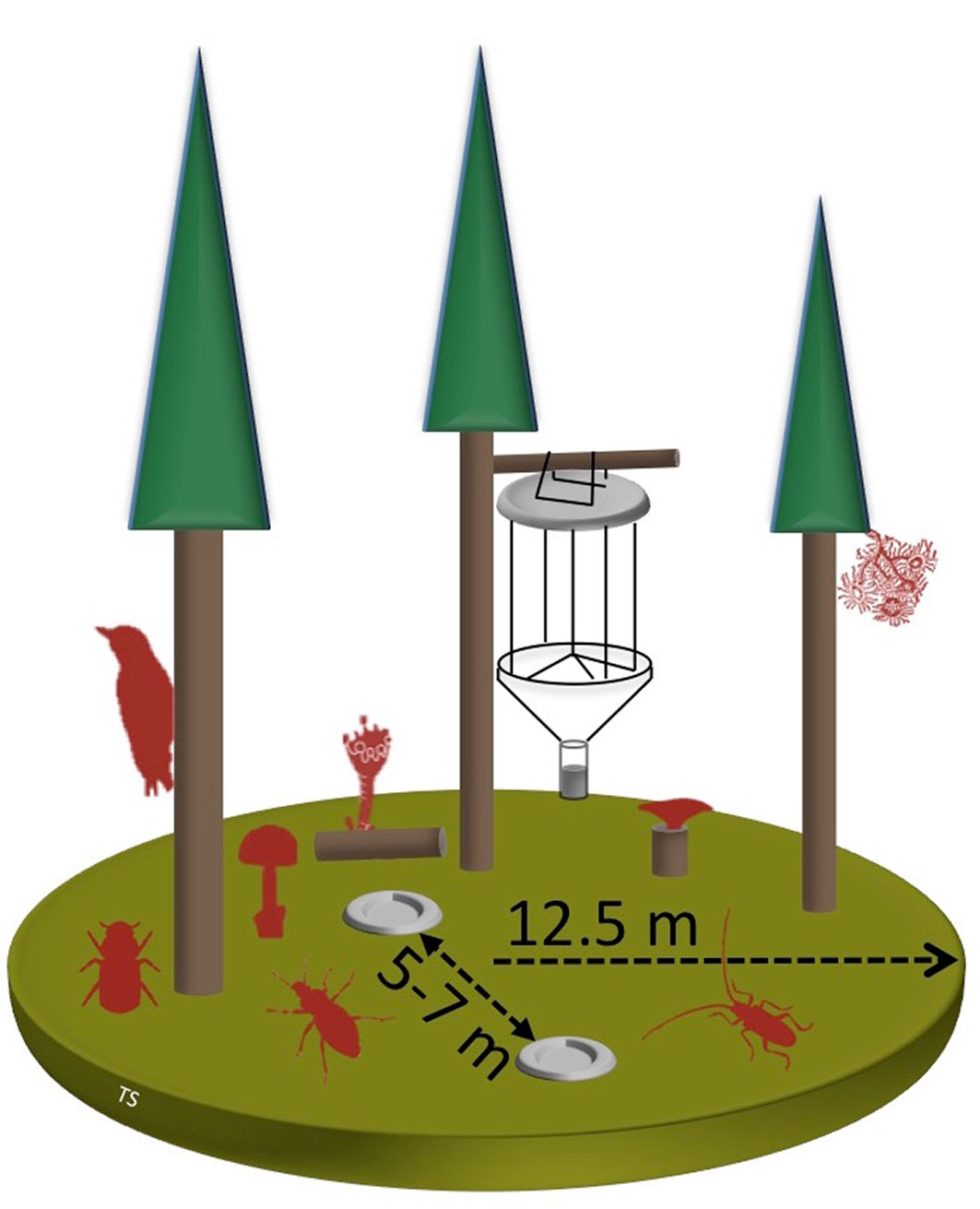
Between 2009 and 2011, we surveyed the occurrence of species belonging to eight taxa at the center of the 12.5 m radius circular plots. The eight taxa surveyed were: (a) deadwood macrofungi; (b) soil macrofungi; (c) epiphytic lichens; (d) deadwood lichens; (e) bark beetles (Scolytinae); (f) longhorn beetles (Cerambycidae); (g) ground beetles (Carabidae); and (h) bird species. Further details on the sampling units and recording methods used for stand structure are available in Sitzia et al. ([42]).
In 2009 and 2010, the occurrence of fruiting bodies of deadwood and soil macrofungi in each plot were recorded in August, September, and October. Each survey was carried out during a similar time-span to improve data comparability between plots. Species that were not directly identified in the field were collected for later identification in the laboratory, and specimens were stored in permanent collections.
The occurrence of all epiphytic lichens present from the tree-base up to 2 m height were assessed in 2009 for all individual silver fir, Norway spruce, and beech trees. Deadwood lichens were surveyed on all stumps, snags, and logs occurring in each plot (see [25] for further details).
Two different traps were used to assess the abundance of beetle species in the plot: two pitfall traps for ground beetles and one window trap for longhorn and bark beetles. Traps were inspected every 15 days between May and mid-October 2009 (see [44] for further details).
Birds were surveyed twice per year during the reproductive season (end-May and June) between sunrise and 11 a.m. Each survey consisted of a 10-minute point count, preceded by 2 minutes of silence to offset the disturbance effect due to the arrival of the surveyor. All bird species which were visually or acoustically detected were noted during the point count.
Species nomenclature followed Padovan ([31]), Nimis & Martellos ([28]), Sitzia et al. ([44]), and Fracasso et al. ([14]) for macrofungi, lichens, beetles, and birds, respectively.
Statistical analysis
Analysis to test the effect of forest management on species diversity was carried out at the plot-level for (a) mean species richness and (b) species composition (presence-absence data) for each taxon. Differences in mean species richness were tested using a Fisher-Pitman permutation test ([5]), and significant findings were further tested using a Monte Carlo randomization test on means, based on 9999 combinations ([27], [50]). Principal coordinates analysis based on a Sørensen’s dissimilarity matrix and a permutational multivariate analysis of variance (PERMANOVA based on 9999 permutations - [2]) were used to test for differences in species composition between the two forest management types. All analyses were performed using the R statistical software version 3.0.2 ([36]), with the Fisher-Pitman permutation test performed using the “oneway_test” function in the “coin” package ([49]); and the PERMANOVA analysis conducted using the “adonis” function in the “vegan” package ([30]).
Results
A total of 34 species of deadwood macrofungi, 78 soil macrofungi, 56 epiphytic lichens, 22 deadwood lichens, 17 bark beetles, 15 longhorn beetles, 19 ground beetles, and 20 birds were found in the two test watersheds (see Tab. S1 in Supplementary material for the original data used to perform the analyses). The total species richness of the 10 sampling plots for each forest watershed (γ-diversity) is reported in Tab. 2.
Tab. 2 - Total number of species in the 10 sampling plots from each forest watershed.
| Taxon | Non-intensively managed | Abandoned |
|---|---|---|
| Deadwood macrofungi | 38 | 62 |
| Soil macrofungi | 25 | 15 |
| Epiphytic lichens | 44 | 39 |
| Deadwood lichens | 16 | 22 |
| Bark beetles | 12 | 17 |
| Longhorn beetles | 6 | 13 |
| Ground beetles | 16 | 14 |
| Birds | 16 | 16 |
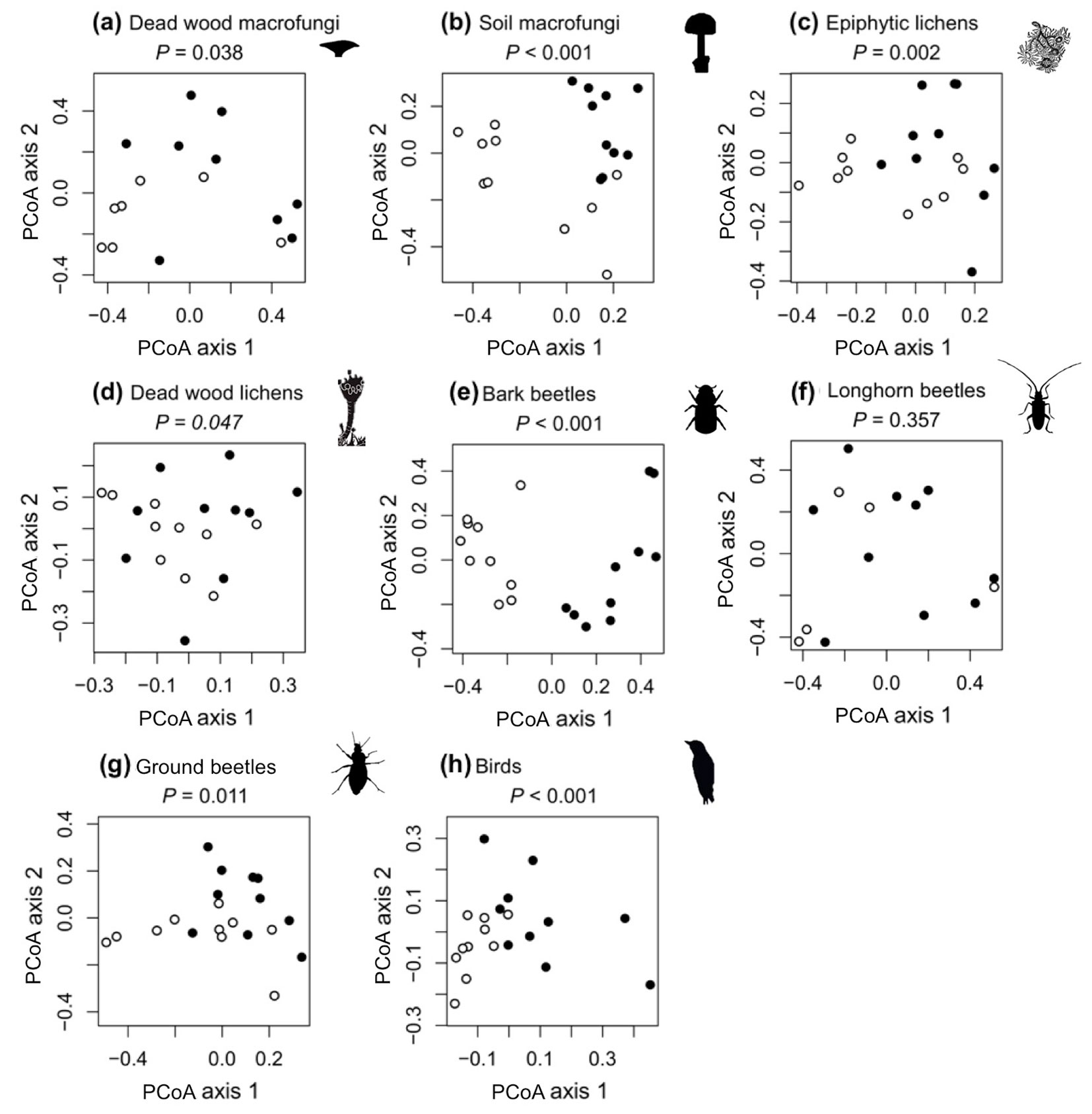
The mean species richness for four taxa (soil macrofungi, deadwood lichens, bark beetles, and longhorn beetles) were significantly higher in the abandoned than in the non-intensively managed forests (Fig. 2). No significant difference in mean species richness was found between the two management regimes for deadwood fungi or epiphytic lichens, while mean species richness for ground beetles and birds was higher in the non-intensively managed than in the abandoned forest. These findings indicate that differences in individual mean species richness do not always reflect that of total species richness (Tab. 2). However, significant differences in species composition were found between the abandoned and non-intensively managed forests for all taxa, with the exception of longhorn beetles (Fig. 3).
Fig. 2 - Mean number of (a) deadwood macrofungi, (b) soil macrofungi, (c) epiphytic lichens, (d) deadwood lichens, (e) bark beetles, (f) longhorn beetles, (g) ground beetles, and (h) bird species in relation to forest management (A: abandoned; M: non-intensively managed). Error bars represent the standard error. The plots report the P-values of permutation based statistical test used to test differences on mean species richness between the two forest management types.
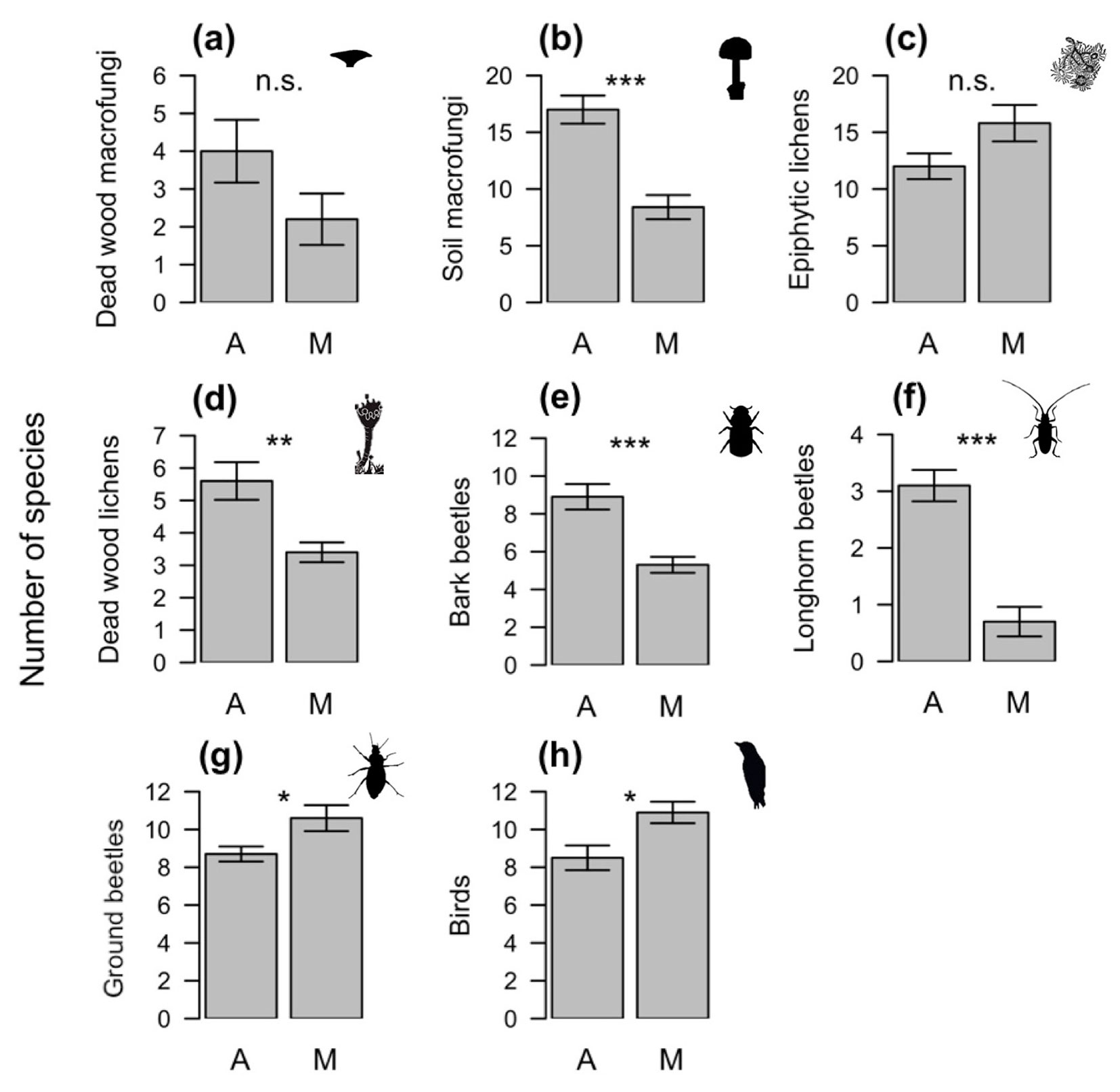
Fig. 3 - Principle coordinates analysis (Sørensen’s dissimilarity matrix) of (a) deadwood macrofungi, (b) soil macrofungi, (c) epiphytic lichen, (d) deadwood lichen, (e) bark beetle, (f) longhorn beetle, (g) ground beetle, and (h) bird community composition. Each point represents the composition of a sampling plot (filled dots: abandoned forests; open dots: non intensively managed forests). The plots report the P-values of a PERMANOVA used to test differences in composition between the two types of forest management types. Sites are plotted only when at least one species was recorded.
Discussion
The results of this study reveal contrasting diversity patterns between the abandoned and non-intensively managed forests for eight different taxa, which encompass a wide range of substrates and resource usage. These findings support the hypothesis that the biodiversity impacts of conversion from intensive forest management to forest abandonment may not be as striking and consistent across taxa as the conversion from intensive to non-intensive forestry ([32]). This may be a reflection of the fact that some species are favored by intermediate disturbance regimes ([38]).
The four taxa whose mean species richness was found to be higher in abandoned forests are mainly related to deadwood (i.e., lichens, bark and longhorn beetles - [25], [44]). This substrate type tends to be more abundant and diverse in abandoned forests, as these sites tend to accumulate higher amounts of logs and snags that improve the range of micro-habitats for organisms colonization. By contrast, in managed forests tree stumps are the only type of deadwood that is typically available ([29], [41]). The amount and diversity of deadwood in abandoned forests are therefore likely to enhance the establishment of less common communities, which are not likely to be found in forests managed solely for timber production ([24], [39]). A consistent pattern was observed in this study between lichens and bark beetles, whereas longhorn beetles showed a different pattern, in which communities similar in composition were progressively enriched by a species accumulation process, coupled with the progressive accumulation of different types of deadwood over the time since abandonment ([44]).
Higher biodiversity was observed for both birds and ground beetles in the non-intensively managed forests compared to the abandoned forests. For birds, this may reflect a negative effect on biodiversity due to canopy closure ([16]) associated with forestry abandonment. Moreover, forestry abandonment promotes the spontaneous reforestation of adjacent open lands and the development of forest gaps, which provide fundamental food and nesting resources for many bird species ([43], [7]). For ground beetles communities, our results agree with previous studies reporting that non-intensive forestry is compatible with high beetle biodiversity ([47]), which may be due to the lower soil moisture levels than in abandoned forests ([44]). As it was found for other taxa in these study, the differences in mean species richness of ground beetles were not associated with differences in species composition, indicating that compositionally similar communities may evolve under different management regimes.
Interestingly, the mean species richness of epiphytic lichens and deadwood fungi did not significantly differ between non-intensively managed and abandoned forests. On the contrary, differences in species composition were detected for both taxa. For lichens, these differences may be due to gradual canopy closure, which would enhance more shade-tolerant species ([42], [25]). Increasing tree age would also be beneficial for lichens, due to an increase in substrate stability and availability, and changes in the chemical-physical features of the bark ([12]). As with the other deadwood related organisms considered in this study, deadwood fungi showed differences in species composition that are likely related to the greater availability of different types of dead wood in abandoned forests, which enhances the establishment of species that cannot be found on stumps in productively managed forests ([1], [34]).
We stress that the differences in diversity observed in mean species richness did not always reflect differences of total species richness. This suggests that further research and sampling is needed to elucidate the variability of species richness at the stand scale, rather than at the plot scale.
Conclusions
The contrasting diversity patterns found in this study between the eight taxa in the abandoned and non-intensively managed forests suggests that management abandonment cannot be generalized as a best practice for biodiversity conservation in mountain forests of the Alps. Rather, these findings support the hypothesis that enhancing forest landscape heterogeneity through the creation of a mosaic of abandoned and non-intensively managed forests could better fulfill the requirements of ecologically different taxa. Such an approach may enhance biodiversity conservation in forest landscapes of the Alps, at least in terms of species richness and composition.
Our results are based on a case study alone and do not allow to make a general inference on the effects of a given management regime. However, our findings can contribute to the formulation of new research hypotheses and to provide valuable data for meta-analyses, which is an increasingly used tool to explore general patterns in ecology ([10]). Moreover, the results of this study can assist in the scientific assessment of adverse effects of forest management ([45]), forest ecosystem functions mapping ([53]), and diagnostic species selection ([8]) in forest habitats. To further examine the hypotheses and findings of this study, more multi-taxon investigations are needed at a variety of landscapes. Such studies should address: (a) a higher replication level, ensuring a more robust evaluation of the effect of the management regime and the exclusion of other possible factors not considered in our study (e.g., historical species distribution and landscape legacy); (b) sites abandoned for longer than 50 years, to allow for an evaluation of the consistency of the effects of abandonment found in our study across a longer time span; (c) turnover and nestedness of compositional heterogeneity (β-diversity); and (d) an experimental approach which can better elucidate the causal relationships between management, structural features, and multi-taxon biodiversity.
Acknowledgements
M.E. Olson is acknowledged for his helpful comments on a previous version of the manuscript. This project was supported by the Italian Ministry of Agricultural, Food and Forestry Policies, State Forestry Corps (Project Managers: A. Andrighetti and D. Campedel), research agreement No. 767/ 2008 (TS, Principal Investigator). We thank M. Varaschin for providing assistance to the bird survey. TC was funded by the Dept. TESAF, University of Padova (PhD grant, 29th cycle). TS designed the methods, carried out surveys, coordinated the research team, developed and wrote the manuscript, TC played a primary role in the manuscript developing, writing and polishing, MD performed the statistical analysis, MC carried out the bird survey, EG and MS carried out the beetle surveys and coordinated the project, FP carried out the surveys of macrofungi, JN carried out the lichen surveys, played a major role in developing and writing the manuscript and collaborated to the sampling design. D. Hawtree performed the language revision of the manuscript.
References
Gscholar
Gscholar
CrossRef | Gscholar
Online | Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
CrossRef | Gscholar
Online | Gscholar
CrossRef | Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Thomas Campagnaro
Matteo Dainese
Department of Land, Environment, Agriculture and Forestry, Università degli Studi di Padova, v.le dell’Università 16, I-35020 Legnaro, Padova (Italy)
Cassol & Scariot Forest and Wildlife Professionals, v. Stadio 18, I-32036 Sedico, Belluno (Italy)
Enzo Gatti
Monica Sommacal
Raggruppamento Carabinieri Biodiversità - Reparto Biodiversità di Belluno, v. Gregorio XVI 8, I-32100 Belluno (Italy)
Mycologist, v. Mier 73, I-32100 Belluno (Italy)
Department of Biological, Geological and Environmental Sciences, University of Bologna, v. Irnerio 42, I-40126, Bologna (Italy)
Corresponding author
Paper Info
Citation
Sitzia T, Campagnaro T, Dainese M, Cassol M, Cortivo MD, Gatti E, Padovan F, Sommacal M, Nascimbene J (2017). Contrasting multi-taxa diversity patterns between abandoned and non-intensively managed forests in the southern Dolomites. iForest 10: 845-850. - doi: 10.3832/ifor2181-010
Academic Editor
Matteo Garbarino
Paper history
Received: Jul 28, 2016
Accepted: Jun 30, 2017
First online: Oct 26, 2017
Publication Date: Oct 31, 2017
Publication Time: 3.93 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2017
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 51189
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 42381
Abstract Page Views: 3663
PDF Downloads: 4019
Citation/Reference Downloads: 18
XML Downloads: 1108
Web Metrics
Days since publication: 3160
Overall contacts: 51189
Avg. contacts per week: 113.39
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2017): 16
Average cites per year: 1.78
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Short Communications
Towards cost-effective indicators to maintain Natura 2000 sites in favourable conservation status. Preliminary results from Cansiglio and New Forest
vol. 1, pp. 75-80 (online: 28 February 2008)
Research Articles
Integrating conservation objectives into forest management: coppice management and forest habitats in Natura 2000 sites
vol. 9, pp. 560-568 (online: 12 May 2016)
Research Articles
Effects of different silvicultural measures on plant diversity - the case of the Illyrian Fagus sylvatica habitat type (Natura 2000)
vol. 9, pp. 318-324 (online: 22 October 2015)
Research Articles
Biodiversity conservation and wood production in a Natura 2000 Mediterranean forest. A trade-off evaluation focused on the occurrence of microhabitats
vol. 12, pp. 76-84 (online: 24 January 2019)
Book Reviews
National forest inventories: contributions to forest biodiversity assessments (2010)
vol. 4, pp. 250-251 (online: 05 November 2011)
Review Papers
The forest biodiversity artery: towards forest management for saproxylic conservation
vol. 9, pp. 205-216 (online: 26 October 2015)
Research Articles
Wildfire incidence in Spanish Natura 2000 sites is largely explained by forest land cover
vol. 19, pp. 195-200 (online: 04 June 2026)
Research Articles
The effects of forest management on biodiversity in the Czech Republic: an overview of biologists’ opinions
vol. 15, pp. 187-196 (online: 19 May 2022)
Short Communications
An approach to measuring biodiversity and its use in analysing the effect of nitrogen deposition on woodland butterfly populations in the Netherlands
vol. 2, pp. 46-48 (online: 21 January 2009)
Research Articles
Investigating the effect of selective logging on tree biodiversity and structure of the tropical forests of Papua New Guinea
vol. 9, pp. 475-482 (online: 25 January 2016)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







