
Assessment of hedge stand types as determinants of woody species richness in rural field margins
iForest - Biogeosciences and Forestry, Volume 6, Issue 4, Pages 201-208 (2013)
doi: https://doi.org/10.3832/ifor0749-006
Published: May 27, 2013 - Copyright © 2013 SISEF
Research Articles
Abstract
The woody component along field edges often provides the only permanent elements of structural and biological diversity in landscapes that have lost much of their naturalness to agriculture and urban use. The main aims of this study were (i) to investigate how four hedge stand types, i.e., systems of management distinguished on the basis of the natural and managed growth forms of trees and shrubs (low single-storied, high single-storied, two-storied and multi-storied), differ in their biometric parameters and (ii) to identify the primary management-related determinants of native woody species richness. The study used a large dataset of hedgerows (n=538) sampled in seven sites in Northern Italy. The four hedge stand types exhibited highly significant differences in size and biometric parameters. The multi-storied hedges, associated with the highest stand structural diversity index values, supported the highest number of tree species, followed by high single-storied hedges. The low single-storied hedge stand type contained the lowest number of species. We found a positive effect of hedge area as well as a marginal negative effect of basal area on native woody species richness. The management implications addressed by our study include conversion of single-storied into two- or multi-storied types, increasing hedge size and controlling hedge stand basal area.
Keywords
Agricultural Landscape, Basal Area, Biodiversity, Hedgerow, Structure, Tree Growth Form, Management, Size
Introduction
The tree and shrub layers of rural hedges often provide the only semi-permanent element of structural and biological diversity in landscapes that have lost much of their naturalness to agriculture and urban use ([39], [11]). These layers are composed of woody species maintained or planted by farmers routinely or in response to regulatory directives ([57], [12]). Hedges with a high diversity of woody species are expected to support a great abundance and diversity of associated organisms such as birds ([29], [24], [38]) and invertebrates ([32]). However, woody species richness is influenced by many factors among which hedgerow structure is of great importance ([25], [63], [57], [23], [8], [16], [17], [13]). Hedgerow structure has been defined in many ways, including as: (i) a combination of different biometric values ([57], [2]); (ii) the percentage cover of the different vegetation layers ([4]); (iii) a combination of these two sets of variables ([37], [21], [20], [23], [54]); and (iv) all of the above, combined with size and other physical measures ([16], [13], [52], [62]). However, despite the number of structure classification schemes, little attention has been paid to the significance of stand biometric differences among the structure types, which are always recognized through visual estimation in the field and hence highly subject to error. The present paper introduces and validates, through a post-analysis of biometric differences among the groups, a criterion for classifying rural hedge stands based on the relative contribution of trees, coppice, pollards and shrubs to the hedgerow. Based on this criterion, primary management-related determinants of native woody species richness are identified. Specifically, a large dataset was used, including data from more than 500 hedges located in the Padua-Venetian intensive peri-urban agricultural landscapes. The use of this shorthand recording system of hedge stand types, which incorporates biometric values and woody species growth form categories, increases the applicability of the existing hedgerow structure classification scheme to planning and management.
Materials and methods
Study area
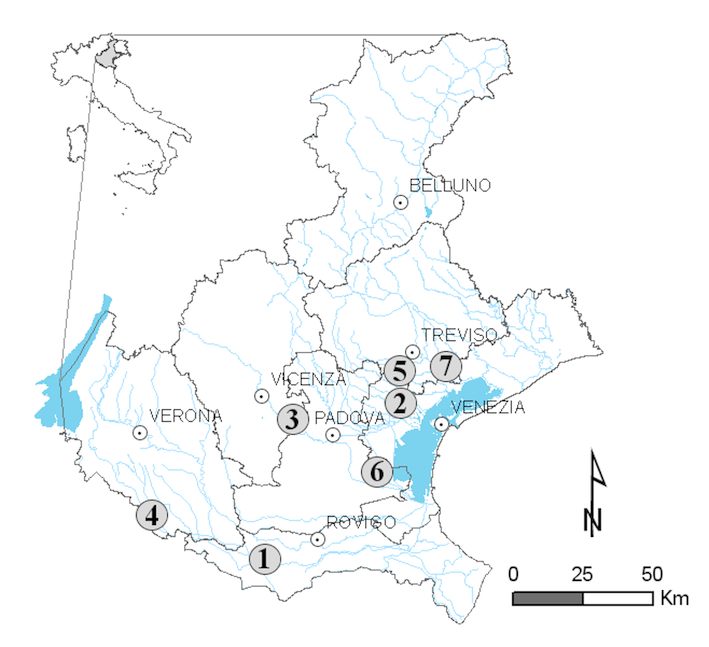
The Padua-Venetian plain is a sub-region of the Po Plain located in the north-eastern part of Italy and occupying an area of approximately 10 372 km2 (Fig. 1). Mean annual precipitation and temperature vary between 700 and 1100 mm and 13 to 13.5 °C, respectively. All the sites are nearly flat, the site with the highest elevation (Montegalda) being located at 63 m a.s.l. The study region is located in the north Mediterranean environmental zone ([35]). The area has been subjected to substantial transformations that have modified the landscape structure, since the Roman centuriation in the 1st century BC up to the industrial settlements in the 1970s ([59]). At present, 49-85% of the region is agricultural and maize, soybean, others cereals and wine grapes are the dominant crops ([48]). Hedgerows were established or modified in different periods, and spontaneous regeneration of plant species is possible where their function is not exclusively ornamental ([64], [55]).
Fig. 1 - Italy, Veneto region and location of the surveyed sites. (1): Canda; (2): Frassinelli; (3): Montegalda; (4): Nogara; (5): Peseggia; (6): Piove di Sacco; (7): Roncade.
Data collection
Site selection criteria were conditioned according to landscape structure, which has been shaped by the aforementioned changes in land use. Seven sites (Fig. 1) were ultimately selected on the basis of differences in agricultural intensity, measured according to mean enterprise Utilized Agricultural Area (UAA) classes as recorded at the municipal level. Data on these classes are easily achievable, and the classification system is linked to the parceling of the rural landscape, which is an important factor in determining the quality of landscape management by the enterprises ([60]).
The UAA classes considered are ([15]):
- high (≥ 10 ha)
- intermediate (3-10 ha)
- low (< 3 ha)
Similarly to the mean UAA, hedgerow density in each site reflects different agricultural landscape assets and land use histories, from the ancient agricultural landscape of Roman centuriation to the intensive agricultural lands reclaimed in the Early and Late Medieval Ages or in more recent times ([58], [34], [49]).
The general definition of a hedgerow is a row of woody plants, including hedge and herbaceous species, that separates adjacent fields ([45], [18]) and is managed to some degree ([3]). In this study we adopted a definition based on several sources ([7], [6]) and define hedgerows using the following criteria (Tab. 1):
Tab. 1 - Thresholds used to define a hedgerow.
- the function is not exclusively ornamental, and spontaneous regeneration of plants is possible;
- the proportion of area in tree and/or shrub cover gaps of 2 m or more must be less than 35%;
- the length of an individual gap must not exceed 20 m; beyond this threshold the gap is considered an opening dividing two different hedgerows;
- the minimum distance between hedgerows must be 20 m;
- the width must not exceed 20 m.
Within each site, all hedgerows fulfilling these requirements were surveyed with the aim of obtaining a total sample of 50-60 km of hedgerow in each UAA class. Therefore, we surveyed from a minimum of 47 to a maximum of 107 sampling units in each site, with a total of 538 hedgerows surveyed (Tab. 2).
Tab. 2 - Surveyed sites.
| Sites | Provinces | Municipalities | UAA classes | Site area (ha) | Sites hedgerow densities (km 100 ha-1) |
No. sampling units |
|---|---|---|---|---|---|---|
| Nogara | Verona | Nogara, Gazzo Veronese, Sorgà | H | 625 | 2.7 | 59 |
| Rovigo | Rovigo | Canda, CastelGuglielmo | H | 4572 | 0.85 | 68 |
| Piove | Padova | Piove di Sacco, Arzegrande, Brugine, Pontelongo | L | 420 | 6.97 | 107 |
| Frassinelli | Venezia | Martellago, Spinea, Venezia | L | 158 | 8.67 | 47 |
| Peseggia | Treviso Venezia |
Zero Branco, Scorzè | L | 175 | 8.14 | 59 |
| Roncade | Treviso | Roncade | I | 645 | 3.79 | 106 |
| Montegalda | Vicenza | Grisignano di Zocco, Montegalda | I | 627 | 3.92 | 92 |
| Total | 7222 | 2.24 | 538 | |||
| Mean (± 95% conf. int.) | 1031 ± 1166 | 5.00 ± 2.19 | 76.86 ± 18.13 | |||
Survey of some hedgerow characteristics involved treating the entire hedge as the sampling unit, while others were measured by subdividing the longitudinal length of the hedge into equal parts. The total number of equal parts was proportional to total hedgerow length (L) as follows: single part (L < 100 m); two parts (100 m ≤ L < 300 m); three parts (300 m ≤ L < 600 m); four parts (L ≥ 600 m). In the center of each part, a sampling subunit of 10 m was surveyed. The values recorded in the subunits were then averaged for the entire hedgerow.
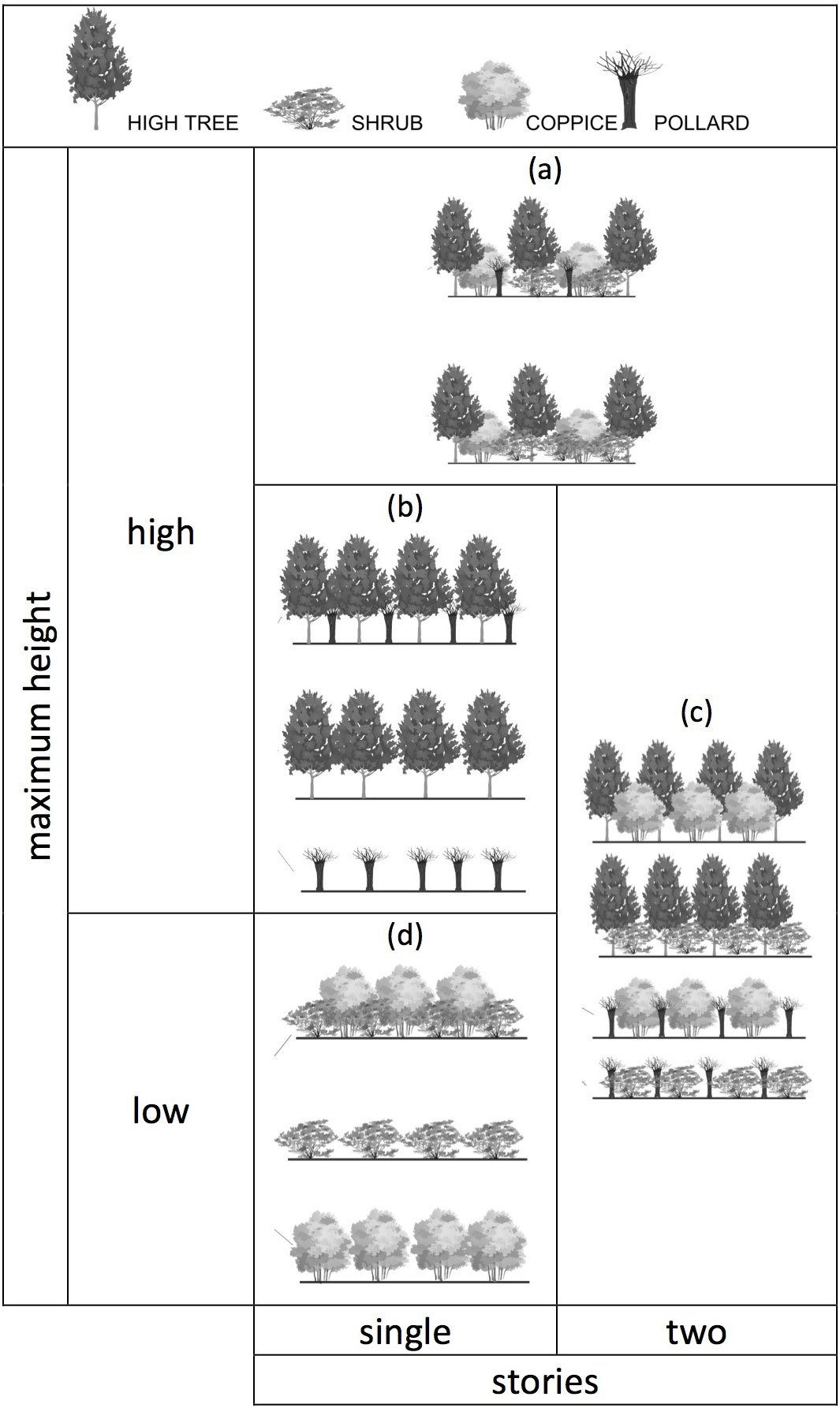
Identification of the following hedge stand types was performed through analysis of the presence of natural and managed growth forms of woodland trees and shrubs (sensu [41], pp. 224 - trees, coppice, pollards, shrubs), which shape the hedgerows and contribute to the varying complexity in vertical structure:
- multi-storied (formed by trees, shrubs, coppices, with or without pollards);
- two-storied (formed by pollards and shrubs, pollards and coppices, trees and shrubs or trees and coppices);
- low single-storied (formed by coppices and shrubs, coppices only or shrubs only);
- high single-storied (formed by pollards and trees, pollards only, or trees only).
Shrubs and coppices usually form the low storey, while pollards and trees compose the high storey (Fig. 2). The surveyed characteristics are defined in detail in Tab. 3.
Fig. 2 - The hedge stand types characterised according to a combination of the maximum height of the growth forms (tree, pollard, coppice, shrub) and the number of stories (one, two, multi) present. (a): multi-storied, (b): high single-storied, (c): two-storied, (d): low single-storied (modified from [56])
Tab. 3 - List and definition [unit of measurement] of the surveyed hedge characteristics and those of the woody plants present. (*): Sampling method indicates whether the whole hedgerow was surveyed (W) or the survey was carried out in subunits of 10 m hedgerow length (S); (**): Trees: single-stemmed individuals in the mature canopy and tall subordinate strata; coppice: trees and shrubs that have been cut and allowed to grow from the residual stump; pollards: trees cut more than one meter above ground level and allowed to grow from the residual stump.
| Characteristic | Sampling method* | Definition and units of measurements |
|---|---|---|
| Hedge stand type | W | Classified according to the growth forms (trees, coppice, pollards, shrubs **) of the woody species composing the hedge [multi-storied, two-storied, high single-storied, low single-storied]. |
| Length | W | Linear development of the sampled unit (surveyed hedgerow) [m]. |
| Width | S | Mean of the widths referring to the projection to the ground of the two points of maximum expansion of the marginal crown on the principal perpendicular axis of the hedge [m]. |
| Diameter | S | Dendrometric diameter of stems with DBH ≥ 5 cm in 1 cm classes [cm]. |
| Height | S | Cormometric height of stems with DBH ≥ 5 cm in 1 m classes [m]. |
| Woody species richness | W | Number of woody species, excluding those with stems only woody at the base. |
We calculated the basal area from the diameters of the full range of stems (DBH ≥ 5 cm) sampled in each hedgerow. Hedge area was equal to width × length. Two indexes of stand structural diversity were calculated: Shannon index (SH) and Gini coefficient (GI) of diameter (SHD and GID, respectively) and height (SHH and GIH, respectively) diversity, as follows (eqn. 1):
where pi is the number of stems of (diameter or height) size class i and n is the number of size classes. SH values range from 0 to ln(n). GI is the arithmetic average of the absolute values of the differences between all pairs of individual measurements (eqn. 2):
where Xi are the sizes (diameter or height) sorted from largest to smallest X1 ≤ X2 ≤ ... ≤ Xn. The Gini coefficient has a minimum value of 0 when all measurements are equal and a theoretical maximum of 1.0 in an infinite population in which all measurements but one have a value of 0: the ultimate in inequality ([26]). Height and diameter values are taken from the complete range of stems sampled in a single hedgerow.
Compared with other measures of stand structure diversity, the two stand structural diversity indexes described above offer many advantages, among which are their ease of calculation and wide usage, an important requisite for adoption by practitioners ([26], [53], [27]).
The presence and absence of woody species was determined by surveying each hedgerow and identifying every species classified as tree or shrub after Pignatti ([42]). Alien species were distinguished according to Celesti-Grapow et al. ([14]).
Data analysis
Differences in biometric parameters between hedge stand types were tested using a one-way ANOVA. When model residuals deviated from the normal distribution, response variables were log- or square root- transformed before analysis.
To test the effects of area, hedge stand type and basal area on the number of native woody species present, a linear mixed model was applied. The model included the variables hedge area, stand types and basal area, their interactions as fixed effects, and site as a random effect to account for spatial nestedness in sampling. The potential interaction between hedge area and stand type was examined to determine whether hedge stand type affects the slope of the species-area curve. The potential interaction between hedge stand type and basal area was also examined to determine whether the effect of increasing basal area on native woody species richness varies with hedge stand type. All statistical analyses were performed using the statistical package R ([47]). As suggested by Pinheiro & Bates ([44]), a sequential F-test was used to investigate the main effects and interactions in all models using the lme function in the nlme library ([43]) in R with the restricted maximum likelihood (REML) estimation method.
Results
Biometric parameters differed significantly between the four hedge stand types (Tab. 4). Multi-storied hedges had the highest values of area as well as of both structural diversity (Shannon and Gini) indexes. Moreover, the highest values of stem basal area ha-1 and of the number of stems ha-1 corresponded with the high single-storied and low single-storied hedge stand types, respectively.
Tab. 4 - Mean values and standard deviations (SD) of parameters for each hedge stand type and results of the ANOVA test. (SHD): Shannon Index for tree diameters; (SHH): Shannon Index for tree heights; (GINID): Gini Index for tree diameters; (GINIH): Gini Index for tree heights; (*): p < 0.001.
| Parameter | Low single-storied (n=123) |
High single-storied (n=90) |
Two-storied (n=130) |
Multi-storied (n=195) |
F | ||||
|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | ||
| Length (m) | 152.7 | 109.7 | 149.7 | 120.84 | 147.1 | 121.6 | 214.0 | 158.7 | 12.8* |
| Width (m) | 4.8 | 1.8 | 5.3 | 1.7 | 5.5 | 1.6 | 6.5 | 2.3 | 21.4* |
| Area (m2) | 754.9 | 697.0 | 795.4 | 687.0 | 821.8 | 737.6 | 1358.1 | 1054.0 | 24.2* |
| Basal area (m2 ha-1) | 22.7 | 31.2 | 55.5 | 89.7 | 38.8 | 49.5 | 35.5 | 31.9 | 12.2* |
| No. stems ha-1 | 4268 | 4439 | 1814 | 1638 | 2464 | 3039 | 2278 | 1861 | 19.3* |
| SHD | 1.37 | 0.60 | 1.66 | 0.58 | 1.51 | 0.63 | 1.97 | 0.56 | 31.1* |
| SHH | 0.99 | 0.55 | 1.03 | 0.55 | 1.19 | 0.58 | 1.49 | 0.50 | 28.3* |
| GINID | 0.09 | 0.07 | 0.08 | 0.08 | 0.12 | 0.08 | 0.13 | 0.05 | 17.6* |
| GINIH | 0.15 | 0.1 | 0.18 | 0.10 | 0.21 | 0.13 | 0.28 | 0.12 | 35.6* |
The most common woody species (those present in ≥ 20% of sampled hedgerows) were: Salix alba L., Cornus sanguinea L., Platanus hybrida Bot., Sambucus nigra L., Rubus ulmifolius Schott, Robinia pseudoacacia L., Acer campestre L., Ulmus minor Miller, Morus alba L., Populus canadensis L., Humulus lupulus L., Hedera helix L. and Corylus avellana L. A complete list of species is reported in the Appendix 1.
Native and total woody species rich- ness were strongly correlated (r = 0.91, p < 0.001). Aliens represented more than one third of the observed woody species (44 out of 122). Alien richness was moderately correlated with native richness (r = 0.41, p < 0.001). Most of the aliens were neophytes, three of which (Robinia pseudoacacia L., Parthenocissus quinquefolia (L.) Planchon, Ailanthus altissima (Miller) Swingle) are considered by Celesti-Grapow et al. ([14]) to be invasive in the Veneto Region.
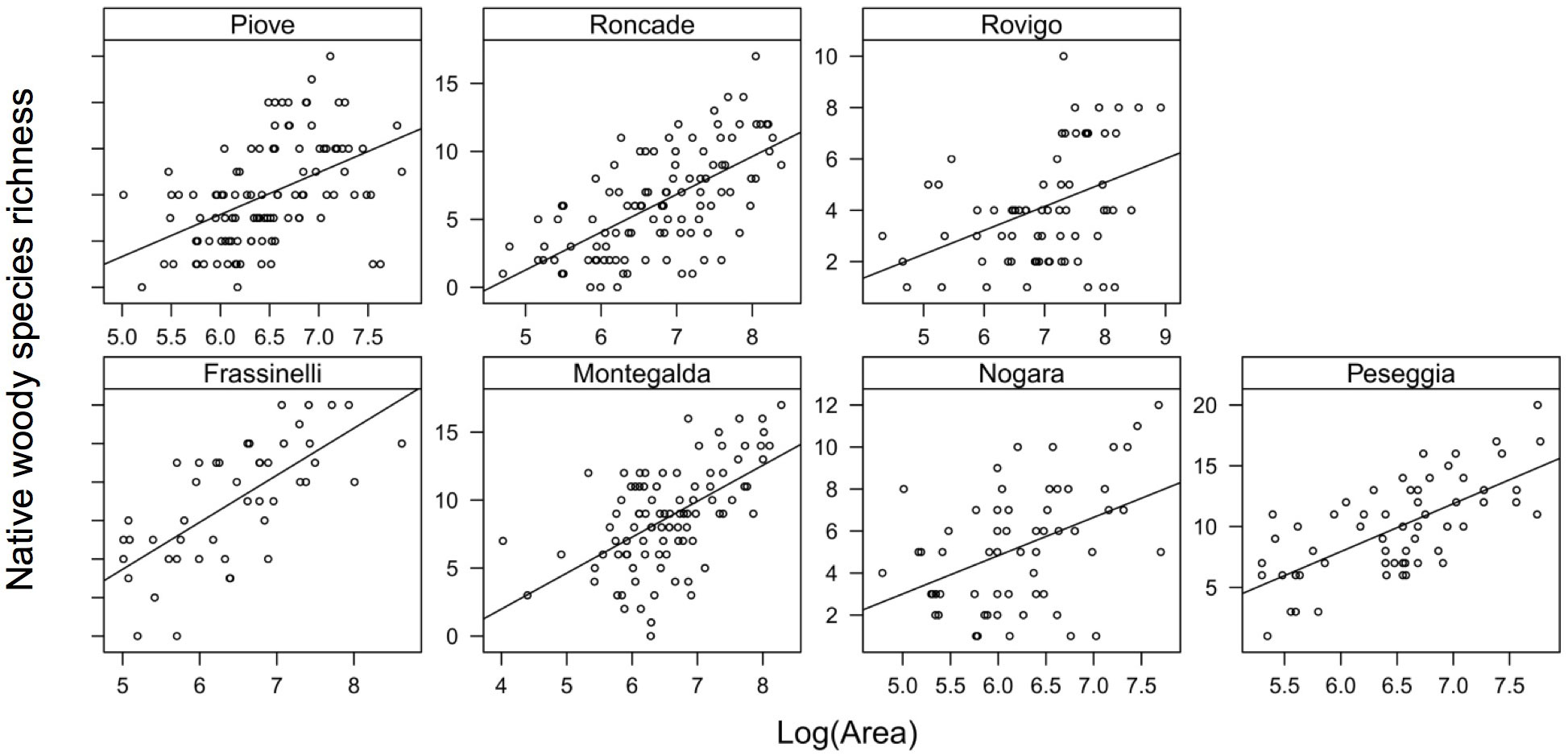
The results of the mixed models (Tab. 5) showed a highly significant effect of hedge area (p<0.001) and stand type (p<0.001) on native woody species richness, as well as a marginal negative effect of basal area (p=0.034). Multi-storied stand types supported the highest number of species followed by two-storied hedgerows. High single-storied stand types contained the lowest number of species (Fig. 3). Interactions between hedge area and stand type and between basal area and stand type were not statistically significant. The slope of the relationship between hedge area and native species richness was positive (Fig. 4) and did not vary with respect to the stand type (Tab. 5).
Tab. 5 - Result from the linear mixed models testing the relationship between native woody species richness and hedge stand type, area, and basal area in the seven sites. No interaction between the factors remained in the model. Site was included as a random factor.
| Parameters | df | F | P |
|---|---|---|---|
| Log (Area) | 1, 526 | 284.74 | <0.001 |
| Hedge stand types | 3, 526 | 38.358 | <0.001 |
| Log (Basal area) | 1, 526 | 4.518 | 0.034 |
| Hedge stand types x Log (Area) | - | - | - |
| Hedge stand types x Basal area | - | - | - |
Fig. 3 - Effect of the four hedge stand types on native woody species richness. White circles within each box represent the mean, while the solid black line indicates the median.
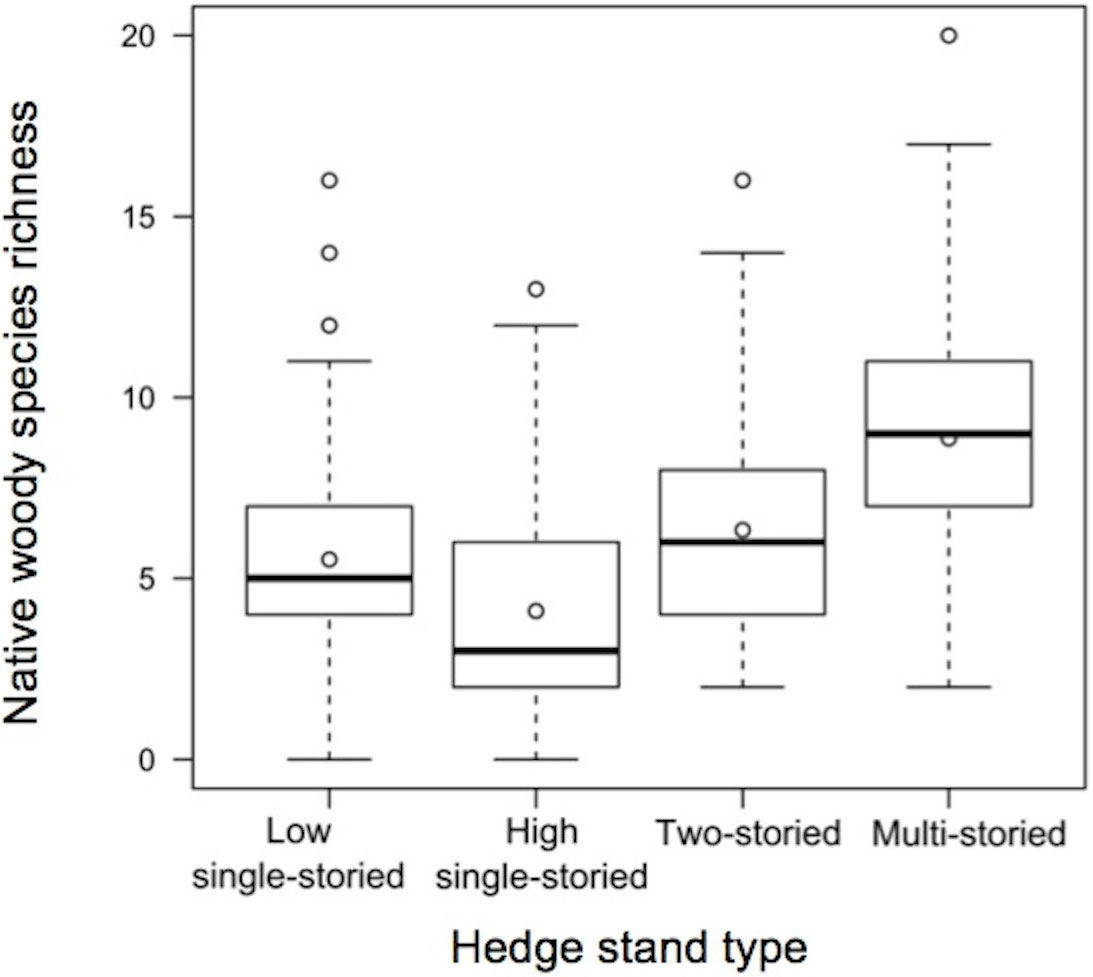
Fig. 4 - Effect of area on native woody species richness. Each plot represents one sampling site (see Tab. 1).
Discussion
Assessment of hedge stand types
Each hedge stand type considered in our study exhibited a relationship with the hedgerow biometric attributes assessed. Native woody species richness was influenced by hedge stand type, area and basal area. As confirmed by the differences in the diversity indexes, basal area and tree density among the four hedge stand types, the presence of several or diverse growth forms guarantees per se higher vertical stratification. Indeed, management practices are typically characterized by cyclic cuttings along the hedgerow, whose frequency and intensity are linked to customs more than to rules, resulting in differences in both vertical tree layer development and values in structural variables. The highest values of the two stand structural diversity indexes are associated with the multi-storied system in which the presence of a multi-layered canopy is associated with a large range of stem diameters and heights and a complex horizontal arrangement of stems ([33]). Similarly, the highest basal area values were found in the high single-storied hedge stand type.
As well documented for forests ([46], [9]), ecosystems including stands with a variety of structural components are considered likely to maintain a variety of resources and species utilizing these resources. Thus, the positive correlation between elements of biodiversity and measures of stand structural complexity observed in other ecosystems can be extended to hedgerow systems, where the more complex vertical arrangement of tree canopy found in the multi-storied hedge stand type supports a greater variety of woody species. These results are consistent with those of Franklin et al. ([19]), who found that species composition and abundance can be indicators of canopy layering. Both high and low single-storied hedgerows contain fewer woody species than do hedgerows with more complex vertical structure, as reported in similar studies conducted in forest stands (e.g., [36]). Microclimatic conditions, the intensity and quality of light, resource availability and other ecological variables can explain the relationship between tree canopy structure and species richness (e.g., [40]). Our results support and extend previous findings describing the relationship between woody species richness and a variety of local hedgerow attributes ([63], [57], [23], [16], [13]) also to the spatial arrangement of tree and shrub growth forms. Most notably, the present study indicates that the identification of hedge stand types, obtained through a visual survey, may allow estimation of native woody species richness across a wide range of hedgerow network densities and landscape types, favoring interaction between decision makers, researchers and landscape users ([61]).
Neither the effect of hedgerow area nor that of tree basal area on woody species richness was affected by hedge stand type. Species accumulation with increasing area sampled is a well know phenomenon that also applies to hedgerows (e.g., [16]). Due to the narrowness and homogeneity of hedgerows, we can hypothesize that high basal area values tend to saturate the space faster than in a wider habitat, i.e., a woodland, where the opposite trend can sometimes be observed ([51], [28]).
Management implications
Management implications for native woody species richness conservation stem from the considerations discussed above. With regard to hedge stand types, the conversion of high and low single-storied hedge to two-storied or multi-storied hedgerow should be promoted. This silvicultural practice is common in forest management ([10], [22]) but is seldom applied to hedgerow systems. As Bannister & Watt ([1]) noted, the effects of different hedge-cutting methods have been reviewed ([30]), but current information derives from anecdotes and observational studies rather than from experimental evidence. Acceptable management options may therefore include different cutting regimes for each hedge component, i.e., small branch cycle (3 to 10 years) for shrub species, coppicing (8 to 30 year cycle) for coppicing species and the tree cycle (30 to 150 years) for tall trees. Pollards can be managed with the traditional pollarding technique (see [50]). The pollard cycle lasts 8 to 15 years depending on species and growth rate, ensuring tree survival and allowing periodic sunlight to reach adjacent woody species. These cutting practices are intended to promote varying growth forms of one or more woody species.
Increasing hedgerow area is a recommended practice for enhancing woody species richness. Habitat availability (i.e., hedgerows, woods) at varying spatial scales has been greatly reduced where agricultural intensification has affected agricultural landscapes ([5]). The development of many planning tools proposing hedgerow requalification and planting ([31]) arising from the Agenda 2000 framework has promoted an increase in the area occupied by existing hedgerow systems. Extending hedgerow length to connect two or more hedges is one possible solution. Another way to increase hedgerow area is to extend hedgerow width, intended as maximum canopy width. To achieve this goal, one can retain untrimmed grassy margins along the hedge, allowing spontaneous regeneration of tree species in the margins. Moreover, new grassy strips of variable width could be created around all field boundaries (not just those occupied by existing hedgerows). Finally, thinning to maintain appropriate hedgerow stem density ([50]) can also be recommended. This technique helps to avoid overly high hedgerow basal area, which can negatively influence woody species richness. The aforementioned suggestions can also be applied to enhance the total number of woody species.
Conclusions and further research
This study identified three important hedgerow features, i.e., hedge stand type, area and basal area, that can be easily shaped by appropriate management practices to attain defined conservation targets. Based on the study results, the following practices can be recommended to enhance native woody species richness, the focus of the study: converting high single-storied and low single-storied to multi-storied hedge stand types, increasing hedgerow length and/or width, and controlling basal area (Appendix 1).
Agro-environmental schemes should therefore support these hedgerow management practices to increase woody species richness in the rural landscape. Future work should include silvicultural studies and tests designed to identify optimal thinning and conversion techniques and their optimal spatial distribution across the landscape.
In addition, this study highlighted the frequent presence of two common alien invasive species, Robinia pseudoacacia L. and Ailanthus altissima (Miller) Swingle, suggesting their spread within hedgerows and in adjacent patches of marginal land where agricultural activities have ceased or been temporarily abandoned. Hence, future investigations should consider interactions between the management actions recommended here and the diffusion of these species. The moderate correlation between alien and native species diversity may suggest that the proposed management practices will promote growth of alien as well as native species. However, alien species distribution compared to that of native species is generally more affected by human actions (e.g., planting) thus highlighting the need to consider other explanatory variables for testing for their significance.
The presented hedge stand types embody a variety of traditional and modern management activities. This miscellany of practices indicates the flexible and adaptive character of hedgerow management to the changing landscape of the Po Plain. In the current intensive agricultural landscape, the presence of hedge features (e.g., shredded trees: trees cut at a height exceeding that of coppices but lower than pollards) linked to traditional techniques underscores the importance of cultural traditions for farmers. However, studies are needed to quantify the current significance and value that farmers and landowners confer to such hedgerows in the Po Plain. In addition, further study is needed to describe the ecosystem services that these linear landscape elements provide and the potential influence of different hedge stand types on the provisioning of such services.
Acknowledgements
We are grateful to A. Berardo (Nogara), C. Bernardi (Roncade), E. Cantarello (Frassinelli and Peseggia), L. Milan (Canda), and A. Rizzi (Montegalda and Piove di Sacco) who performed field sampling in the sites indicated in parentheses. We thank D. McCollin for his useful comments and constructive suggestions on earlier versions of this manuscript. TS and GT are also grateful to F. Viola for his valuable guidance and expertise during their academic work. The present research was partially supported with funds provided by the University of Padova (ex 60% 2009 project code 60A08-4001/09 “Analisi ecologica dei sistemi arborei della pianura Padana orientale”) and by the municipality of Piove di Sacco.
References
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Supplementary Material
Authors’ Info
Authors’ Affiliation
G Trentanovi
D Cattaneo
P Semenzato
Department of Land, Environment, Agriculture and Forestry, University of Padova, v.le dell’Università 16, I-35020 Legnaro (PD - Italy)
DAFNAE-Entomology, University of Padova, v.le dell’Università 16, I-35020 Legnaro (PD - Italy)
Corresponding author
Paper Info
Citation
Sitzia T, Trentanovi G, Marini L, Cattaneo D, Semenzato P (2013). Assessment of hedge stand types as determinants of woody species richness in rural field margins. iForest 6: 201-208. - doi: 10.3832/ifor0749-006
Academic Editor
Paola Mairota
Paper history
Received: Aug 27, 2012
Accepted: Mar 05, 2013
First online: May 27, 2013
Publication Date: Aug 01, 2013
Publication Time: 2.77 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2013
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 61122
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 49501
Abstract Page Views: 3999
PDF Downloads: 5713
Citation/Reference Downloads: 37
XML Downloads: 1872
Web Metrics
Days since publication: 4776
Overall contacts: 61122
Avg. contacts per week: 89.58
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2013): 15
Average cites per year: 1.15
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Investigating the effect of selective logging on tree biodiversity and structure of the tropical forests of Papua New Guinea
vol. 9, pp. 475-482 (online: 25 January 2016)
Review Papers
The forest biodiversity artery: towards forest management for saproxylic conservation
vol. 9, pp. 205-216 (online: 26 October 2015)
Research Articles
The effects of forest management on biodiversity in the Czech Republic: an overview of biologists’ opinions
vol. 15, pp. 187-196 (online: 19 May 2022)
Research Articles
Bird response to forest structure and composition and implications for sustainable mountain forest management
vol. 19, pp. 18-27 (online: 11 January 2026)
Technical Advances
Improved estimates of per-plot basal area from angle count inventories
vol. 7, pp. 178-185 (online: 17 February 2014)
Book Reviews
National forest inventories: contributions to forest biodiversity assessments (2010)
vol. 4, pp. 250-251 (online: 05 November 2011)
Research Articles
Consistency among forest structure and biodiversity potential index (IBP): an assessment of stand structural complexity for floodplain poplar woodlands
vol. 18, pp. 335-343 (online: 04 November 2025)
Review Papers
Biodiversity assessment in forests - from genetic diversity to landscape diversity
vol. 2, pp. 1-3 (online: 21 January 2009)
Research Articles
Biodiversity conservation and wood production in a Natura 2000 Mediterranean forest. A trade-off evaluation focused on the occurrence of microhabitats
vol. 12, pp. 76-84 (online: 24 January 2019)
Research Articles
Stand structure and deadwood amount influences saproxylic fungal biodiversity in Mediterranean mountain unmanaged forests
vol. 9, pp. 115-124 (online: 08 September 2015)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







