
Alternative methods of scaling Eucalyptus urophylla trees in forest stands: compatibility and accuracy of volume equations
iForest - Biogeosciences and Forestry, Volume 11, Issue 2, Pages 275-283 (2018)
doi: https://doi.org/10.3832/ifor2155-011
Published: Mar 29, 2018 - Copyright © 2018 SISEF
Research Articles
Abstract
This study developed and tested a procedure that aimed to reduce the cost of forest stand volume estimation. Using a typical 3 x 3 m Eucalyptus urophylla plantation, estimates based on a simplified approach were compared with those of precise but costly reference methods. The simplified approach required measuring the total height and bole diameters up to 2 m high. The volume of the lower part was estimated using Smalian’s formula, while the volume of the upper part was estimated using a simple solid as an approximation. Three typical solids were tested: paraboloid, cone, and neiloid. The approach proposed is non-destructive, because it does not require tree felling, while precise methods are destructive. The operational (traditional) method uses Smalian’s formula to measure bole diameter at short intervals over the whole bole, while the precise, more research-suited (reference) method records water displacement (using a xylometer) to accurately measure the volume of each bole section. The reference and traditional methods, as expected, produced very similar results. The approach proposed, using a paraboloid for the upper part, provided results that were not statistically different to the reference values. The volumes estimated by the proposed approach were used to calibrate the Schumacher-Hall function, and the performance of the model was evaluated using the values obtained by the xylometer.
Keywords
Forest Inventory, Rigorous Scaling, Dendrometric Prototype, Volume Equation, Modeling
Introduction
The quantification of wood volume in forest stands is imperative for the deployment of sustainable forest management plans ([43], [27], [18], [28], [1]). Volume is a variable of great importance for evaluating the growth and yield potential of a forest. It is a basic component of information in any planning process of forest production ([4]); therefore, it is essential to estimate it with a high level of reliability. Biometric scientists have contributed to the development of the methodology for volumetric estimates by developing better and more consistent modeling, as is used today. The first contribution is attributed to Schuberg ([34]), who is considered the first person to introduce the form quotient between the diameters d0.5h and d1.30 to express the bole form of a tree. He applied this concept to the silver fir (Abies alba Mill.), which triggered numerous subsequent research studies on tree forms in Europe. The analytical solution for estimating the form factor as a function of a form quotient qi and a tree height h was first proposed by Schiffel ([32]), as presented in eqn. 1:
where v is tree volume, d is tree diameter at breast height (DBH), h is tree height, ai and βi are coefficients of the model, and q is the proposed form quotient.
Because of its practical applications for various European forest species, it stimulated the development of other formulations and conceptions of volumetric equations. Soon after, the Swedish researcher Höjer presented an analytical form for a forest species ([13]). Later, Prodan ([27]) presented an evolutionary synthesis of form quotients.
The sectioning of tree boles into different geometric bodies has been considered previously (since the first biometric studies), and is an important approach in obtaining their respective volumes ([39], [29], [6], [37]).
Hohenadl ([11]) reported that dominant trees have higher relative increments at the bottom (i.e., in the neiloid segment) to ensure a better balance of trunk weight. He concluded that in these cases the volume is associated with genetic variables, as well as environmental and silvicultural practices.
Larson ([16]) stated that tree form varies depending on edaphic conditions, climate change, the ways in which silvicultural practices in forest stands are conducted, and the sociological position in which each tree grows. It also varies according to the age of the forest stand, either as a natural evolutionary phenomenon, or due to the competitive status of each tree in relation to its closest neighbors.
Row & Guttenberg ([31]) recommend that for better detection of variations in tree stem form at least three distinct sections should be taken.
In the 1960s, several other important studies on tree form were published, including a proposal for the so-called “taper function” ([16], [27], [9], [36], [15]).
It was assumed in the case of sectioned stems that their forms could be represented complementarily as geometrical solids: a neiloid at the base, a paraboloid in the central part, and a cone at the terminal part ([31], [18]). There have even been proposals to use different mathematical functions for each segment. In his study of Picea excelsa, Roiko-Jokela ([30]) retained the three segments, but used a hyperbola to describe the neiloid, a logarithmic function to describe the paraboloid, and a straight line to describe the cone. There are still researchers who separate the stem into more than three sections, but such proposals have increased the difficulty of adjusting the mathematical functions that describe them ([26], [17], [12]).
In forest inventories, the volume of an individual tree is normally obtained by indirect techniques such as volumetric equations, taper functions, and the form factor ([9], [18], [12], [33], [19], [24]), which are derived from direct measurements of the volumes of some trees in the forest stand. Volumetric equations, which are obtained from the adjustment of regression models and are based on existing relationships between volume and easily measurable variables in the field (e.g., tree diameter and height), are the most commonly used procedures to estimate volume ([1], [5]). They have satisfactory efficiency ([4], [33]), and are in current, well-known volume models ([43], [27], [18]).
Of the several methods of measuring tree volume, water displacement (using a xylometer) is the only technique that provides the (true) volume value, and is usually used as a reference in accuracy evaluations of other methods ([22], [2], [19]). However, due to the operational limitations inherent in this method, the (true) volume of individual trees has been estimated with good accuracy by rigorous scaling (measurement of the diameter and height along the bole) using mathematical expressions (e.g., Smalian, Huber, Newton, Hohenadl, and Pressler, among others). This requires taking measurements of felled trees, or the rigorous volume estimation of standing trees using specialized equipment. In Brazil, Smalian’s method has traditionally been used for most forest surveys, by normally defining short sections up to 2 m in height and 1 or 2 m sections for the rest of the trunk, more for the convenience of measurement than for any accuracy reasons ([19]).
Rigorous scaling is normally performed with sample trees felled in plots of the forest inventory or in areas close to them, an activity that is not part of the yearly routine of measurements in the forest inventory. In addition, there are situations in which it is impossible or impractical to obtain scaling data that provide precise volume estimates for the remaining trees in the stand. Therefore, the use of alternative methodologies for scaling in forest inventories could increase the rate of data collection, reduce costs, and minimize technical and operational difficulties ([3]). Preferably, this should be conducted without felling trees (non-destructive methods) using complex equipment to measure only a few bole diameters.
One alternative would be to measure a few diameters of standing trees up to a height that is easily measurable, assuming that the stem from that point to the top can be treated as a single section. In this way, tree scaling would be optimized, because the volume up to an easily measurable height is directly obtained using a traditional tree-scaling method. From that point up, the volume is estimated using a geometrical solid to express the stem form, i.e., using dendrometric prototypes (paraboloid, cone, or neiloid). This methodological proposal is justified, because in commercial plantations, tree boles tend to resemble these geometrical forms. However, the lower part usually has a more irregular form ([20]). Such a configuration using dendrometric prototypes must be conducted with great accuracy, because the lower part accounts for the greatest part of the overall volume ([5]).
Although the volumes of a cone and neiloid can be used to generate a wide variety of shapes, the volume of a paraboloid is the most appropriate for this modeling.
If its base is considered at a height of 2 m for obtaining the volume, then (eqn. 2):
where g2 is the cross-sectional area measured at a height of 2 m on the stem and d2 is the diameter measured at 2 m.
For the paraboloid, the volume and the form factor are defined in eqn. 3 and eqn. 4, respectively:
Where v2 is the volume of the paraboloid, g2 is the cross-sectional area measures at 2 m, h2 is the height of the bole above 2 m, f2 is the form factor of the bole measured above 2 m and d2 is the diameter measured at 2 m.
The development of the paraboloid form depends on f2, as defined in eqn. 4. We initially decided to use Apollonius’s paraboloid (form factor equal to 0.5) to scale Eucalyptus urophylla trees without felling them. In addition, we compared the results obtained by rigorous scaling using Smalian’s method, which is traditionally applied to measure tree volumes in commercial plantations of Eucalyptus in Brazil ([40]), with those obtained using a xylometer. We also evaluated the effects of different scaling methods on the accuracy of volumetric equations adjusted to the forest stand.
Based on the above considerations, we formulated the following hypotheses: (1) the estimated E. urophylla tree volume, as composed of a base that is evaluated using Smalian’s method with detailed measurements taken up to a height of 2 m and an upper part that is evaluated as a basic paraboloid, is similar to that obtained using a xylometer, or the volume obtained by Smalian’s method applied to the entire stem; (2) the volume function calibrated with the values estimated by the simplified approach would approximate very well to the reference (xylometer) estimates.
Material and methods
Study area
The study was conducted in a 7-year-old E. urophylla S. T. Blake plantation spaced at 3 x 3 m in Rio Verde, southwest Goias state, Brazil, which belongs to the agro-industrial cooperative of farmers of that region (COMIGO). The plantation is at an average altitude of 700 m at 18° 00′ 45″ to 18° 01′ 45″ S and 50° 52′ 45″ to 50° 53′ 15″ W. The climate of the region, according to the Köppen classification, is Aw, tropical humid, and is characterized as having two well-defined seasons: dry in the autumn and winter, and wet with heavy rain in the spring and summer. The annual average air temperature ranges from 20 to 25°C, and the rainfall is between 1.200 to 1.500 mm, with an annual average of around 1.300 mm ([38]).
The dominant soils have B Latosol and A moderate horizons and a clay texture, and are classified by EMBRAPA ([46]) as Red Latosols, corresponding to Red Oxisols in the USDA Soil Taxonomy ([42]) and Ferralsols by WRB ([14]).
Data
One hundred and fifty trees in different diameter classes were randomly selected in the population and cut down for the present study. DBHs were measured using a caliper, and ranged from 7.2 to 20.6 cm. The total heights of standing trees were measured using a Vertex hypsometer, and those of felled trees were measured using a tape measure, and ranged from 14.2 to 30.6 m.
After measuring the DBHs and total heights, a rigorous scaling of each tree was performed to obtain its actual volume. This was conducted using three non-destructive methods by scaling standing trees and two destructive methods by cutting trees down.
The scaling of standing trees up to a stem height of 2 m was performed using Smalian’s method and by taking detailed measurements. From that height up, the remaining part of the stem was assumed to include three distinct dendrometric prototypes, i.e., a paraboloid, a cone, and a neiloid (Fig. 1). The diameters of boles with bark were measured at 0.1, 0.3, 0.5, 0.7, 0.9, 1.1, 1.3, and 2.0 m above ground level, and from that height up, at every 1 m up to a stem diameter of 4 cm (commercial). The three non-destructive methods are hereafter referred to as Smalian + Paraboloid, Smalian + Cone, and Smalian + Neiloid.
Fig. 1 - Schematic representation of tree scaling using three non-destructive methods and the traditional Smalian’s method (destructive). The non-destructive methods were Smalian + Paraboloid, Smalian + Cone, and Smalian + Neiloid. (L): length of the section (m); (H): total height (m); (gi): sectional area at height i (m²); (vi): volume of section i (m³); (vParaboloid): volume of paraboloid (m³); (vCone): volume of cone (m³); (vNeiloid): volume of neiloid (m³); (vPonta): tip volume (m³); (v): volume of the tree (m³).
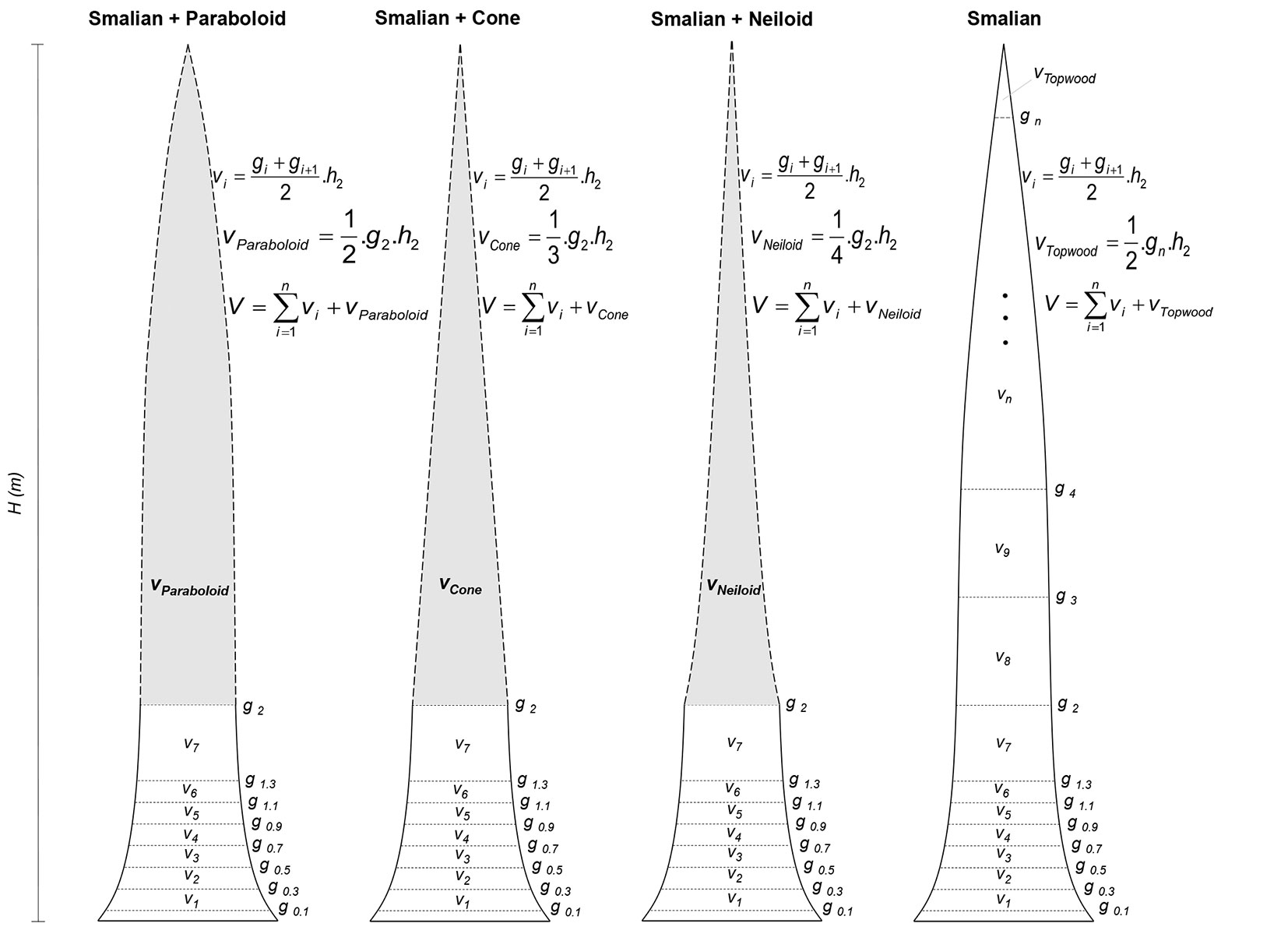
After taking measurements of the standing trees, they were cut at 0.1 m above the ground. The length of the upper part was calculated by taking the difference between the total and commercial heights.
When using Smalian’s method ([19]) on each felled tree, volumes were obtained for all intermediate sections along the tree bole, i.e., between 0.1 m above the ground up to the minimum merchantable diameter (4 cm).
Another destructive method used was the xylometer, by which the volume was obtained by sectioning each tree and immersing the sections (small logs) in a container with water. The water volume displaced corresponded to the true wood volume (Archimedes’ principle). The xylometer used in this study was similar to the one described by Machado et al. ([22]), and was made of two connected drums (total capacity of 200 L) with leveling screws on the bottom and a graduated (dm³) external ruler for measuring the displaced water volume. According to Machado & Figueiredo Filho ([19]), the xylometer method has been used as a reference to evaluate the accuracy of other techniques for obtaining the real volume. It has the advantage of being independent of trunk form, and eliminates some measuring errors, as the diameters and lengths of sections along the stem are not measured.
When comparing the tree-scaling methods (destructive and non-destructive), stump volume (0 to 10 cm) and the upper part’s volume were not included in the evaluation methods. These volumes were not included when using the xylometer because after cutting, the stump and upper part of the tree are usually left in the forest stand.
Evaluation of tree-scaling methods
The efficacy of the different tree-scaling methods in obtaining the volumes of individual trees was evaluated using a Kruskal-Wallis test (α = 0.05), with the volume obtained by the xylometer used as a reference. Five treatments (Xylometer, Smalian, Smalian + Paraboloid, Smalian + Cone, and Smalian + Neiloid) and 150 replications (trees rigorously scaled) were included. The analyses were conducted using the software Sbpank ver. 2.17c ([10]).
The volumes obtained by the different scaling methods were evaluated a posteriori by the aggregate difference (AD), which is the difference between the sum of the volumes obtained by the xylometer and the volumes obtained by the other scaling methods ([23]). We also investigated the distribution of residuals, which indicates under or overestimates when using each of the proposed scaling methods.
Model adjustment and validation
Volume data obtained by the xylometer and the other scaling methods, which were not statistically different, were used to adjust the Schumacher & Hall ([35]) volumetric model to assess the influence of each method on the accuracy of the estimates obtained by the volumetric equations. To achieve this objective, the trees were randomly grouped into two datasets: one for model adjustment using data from each scaling method (100 trees), and the other for model validation (50 trees).
The Schumacher & Hall model was adjusted using Statistic 7.0 software ([44]) with the Levenberg-Marquardt algorithm and 1.000 iterations. The quality of the adjustment was evaluated by the significance of the regression and its parameters (α = 0.05), and by the following measures of precision: coefficient of determination (R²), residual standard error as a percentage (Syx%), and a graphical analysis of the residuals ([7]), in addition to investigating correlations between the observed and estimated values ([25]).
For model validation, a comparison was made of the volumes estimated by each adjusted equation and those obtained by the xylometer using the Kruskal-Wallis test (α = 0.05). Correlations (r) between the values observed (xylometer) and those estimated by the other methods (Syx%x and AD) were assessed, in addition to a graphical analysis of the errors.
Results
Tab. 1 shows that the tree volumes in the E. urophylla stand differed between the different tree-scaling methods. Traditional tree scaling using Smalian’s method (destructive), and scaling using the Smalian + Paraboloid non-destructive method proposed in this study, were statistically similar to the xylometer method (p = 0.8663 and 0.9549, respectively), which is considered a reference for comparison with other methods.
Tab. 1 - Performance of different scaling methods to obtain Eucalyptus urophylla tree volumes. Means followed by the same letter in a column did not significantly differ according to a Kruskal-Wallis test (α = 0.05); (AD): aggregate difference.
| Treatment | n | Volume (m³) | AD (m3) |
AD (%) |
||
|---|---|---|---|---|---|---|
| Mean | Min | Max | ||||
| Xylometer | 150 | 0.2174 a | 0.0314 | 0.4832 | - | - |
| Smalian | 150 | 0.2165 a | 0.0315 | 0.4806 | 0.1306 | 0.40 |
| Smalian + Paraboloid | 150 | 0.2190 a | 0.0294 | 0.4991 | -0.2438 | -0.75 |
| Smalian + Cone | 150 | 0.1577 b | 0.0223 | 0.3548 | 8.9549 | 27.46 |
| Smalian + Neiloid | 150 | 0.1271 c | 0.0188 | 0.2826 | 13.5543 | 41.56 |
However, the average tree volumes as estimated by the other two non-destructive rigorous scaling methods (Smalian + Cone and Smalian + Neiloid) did significantly differ to the mean volumes obtained by the xylometer method (p < 0.0001), the traditional Smalian’s scaling method (p < 0.0001), and the Smalian + Paraboloid method (p < 0.0001). In general, the Smalian + Cone and Smalian + Neiloid methods tended to underestimate the volume obtained by the xylometer method by more than 27%, and significantly differed between themselves (p < 0.0001).
A graphical analysis of the errors associated with the results of the individual tree volumes obtained by each tree-scaling method in relation to the xylometer method showed that the traditional (destructive) Smalian’s technique had the greatest accuracy (-0.64 to 0.82%), despite having a slight tendency to underestimate the volume (Fig. 2a and Fig. 2b).
Fig. 2 - Graphical analysis of errors generated by Eucalyptus urophylla volumes. Different tree-scaling methods were used in relation to the volume obtained by the xylometer. Individual error distribution (a) and histograms of errors (b, c, d, and e).
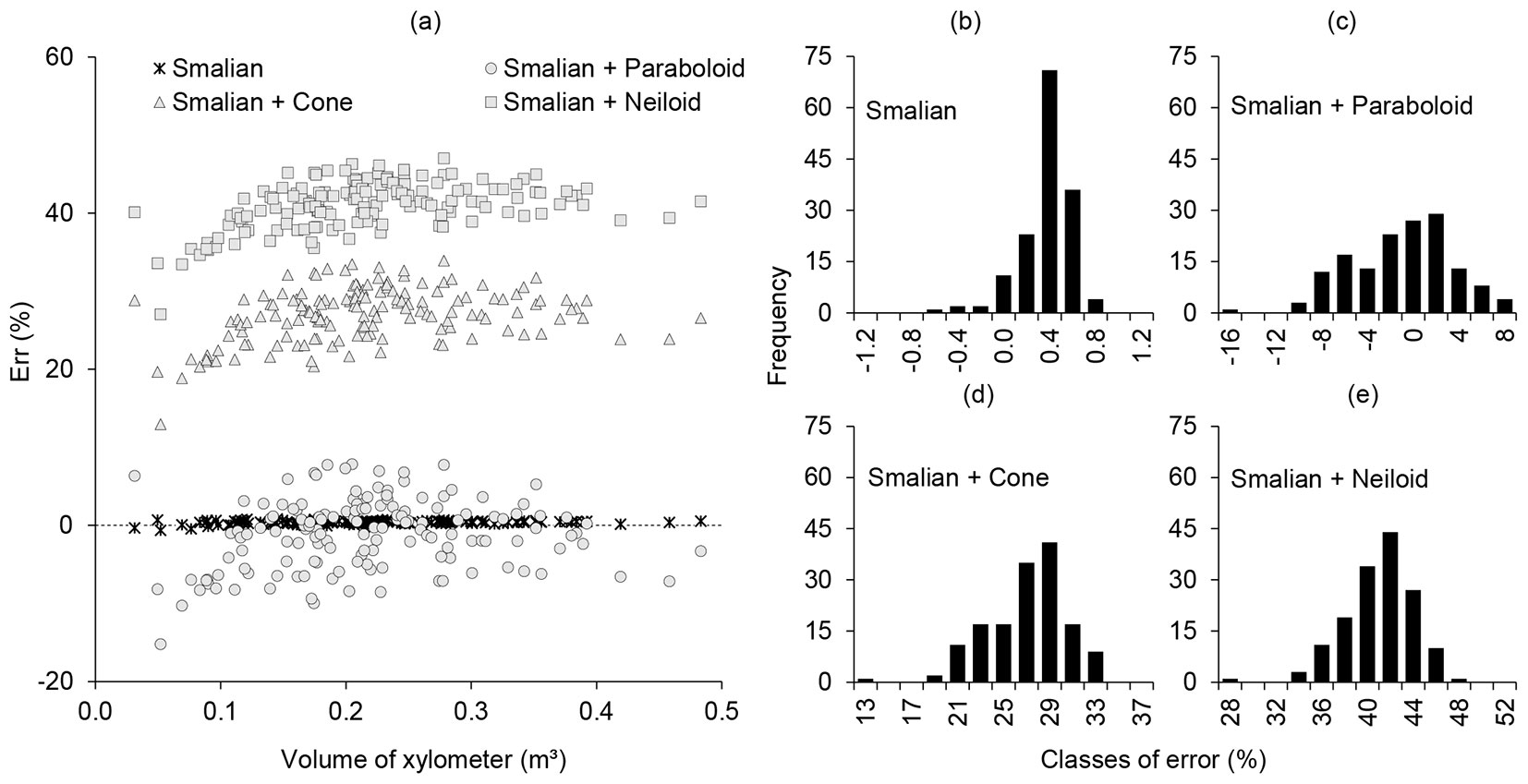
The three non-destructive methods proposed in this study exhibited more expressive dispersion of errors (Fig. 2a). The Smalian + Paraboloid method, with errors varying from -15.21 to 7.83%, had a slight tendency to overestimate the volume (Fig. 2a and Fig. 2c), whereas the Smalian + Cone (Fig. 2a and Fig. 2d) and Smalian + Neiloid methods (Fig. 2a and Fig. 2e) underestimated the tree volumes by 12.97 to 33.93% and 27.06 to 47.01%, respectively.
We applied the volume data obtained by the destructive and non-destructive scaling methods to adjust the volume equations, using the Schumacher & Hall model for each method. The volume equations obtained by the tree scaling methods that were compatible with the xylometer method (Smalian and Smalian + Paraboloid) had similar statistics for adjustment and accuracy, with a relatively high value of R² (>0.97) and a relatively low value of Syx% (<6.65%) (Tab. 2). In addition, they were significant for all of the coefficients of the equations (p < 0.0001). All of the equations predicted strong correlations between the xylometer values and the estimated values (Fig. 3a, Fig. 3b, and Fig. 3c), and provided satisfactory distributions of residuals with errors of between -18.91 and 12.89% (Fig. 3d, Fig. 3e, and Fig. 3f).
Tab. 2 - Volumetric equations with their respective statistics of adjustment, obtained from data applied to different rigorous tree-scaling methods (Xylometer, Smalian, and Smalian + Paraboloid) on Eucalyptus urophylla trees. (V): volume (m³); (DBH): diameter at breast height (cm); (H): total height (m); (R2): coefficient of determination; (Syx%): residual standard error (%).
| Tree-Scaling Method | Volumetric Equation | R² | Syx% |
|---|---|---|---|
| Xylometer | V = 0.000020 · DBH 1.588006 · H 1.539417 | 0.9717 | 6.64 |
| Smalian | V = 0.000022 · DBH 1.608694 · H 1.495780 | 0.9721 | 6.59 |
| Smalian + Paraboloid | V = 0.000019 · DBH 1.756855 · H 1.419518 | 0.9800 | 5.21 |
Fig. 3 - Correlations between observed and estimated volumes and the distribution of residuals from volumetric equations. The following methods were used: xylometer (a and d), Smalian (b and e), and Smalian + Paraboloid (c and f).
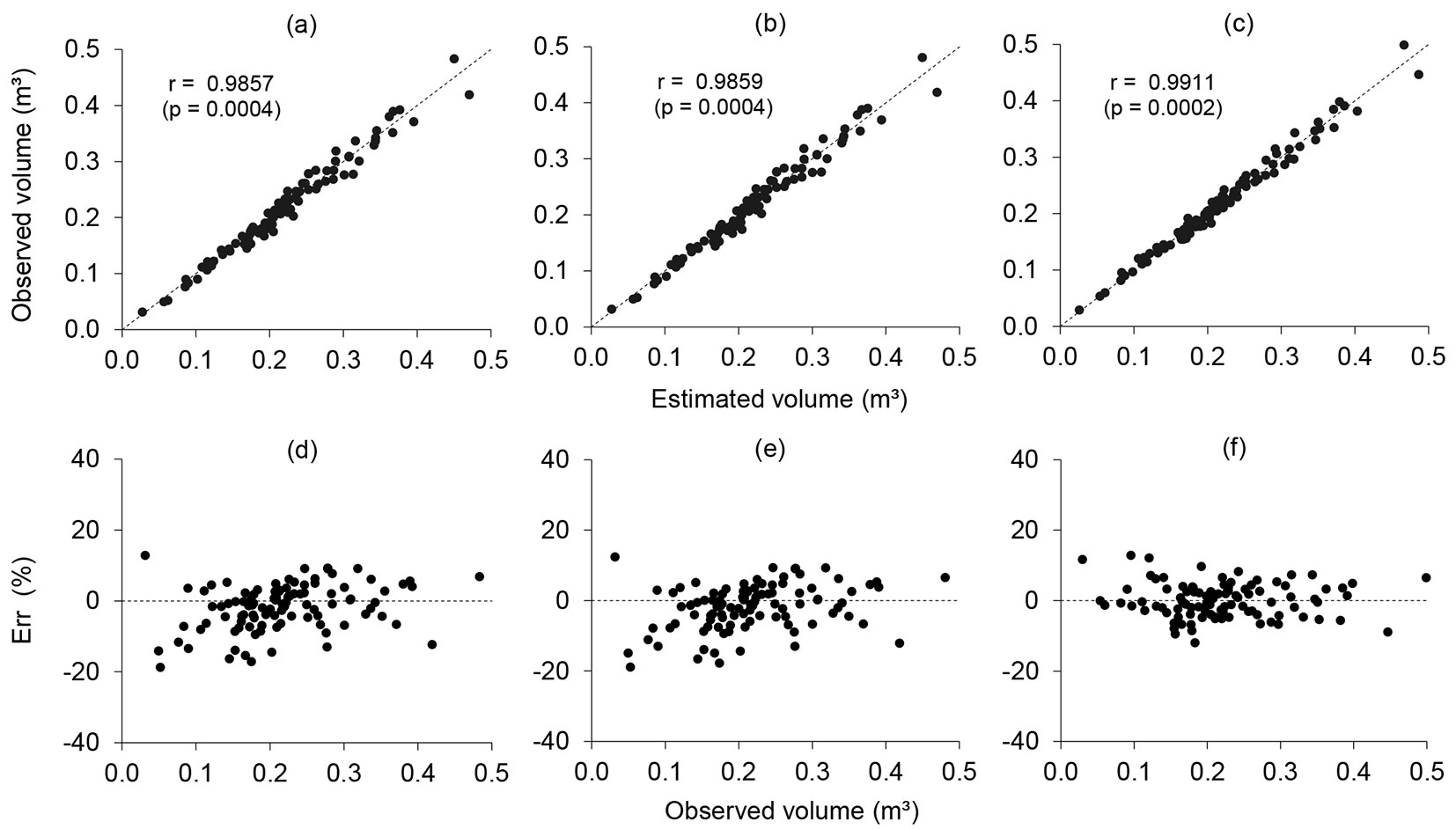
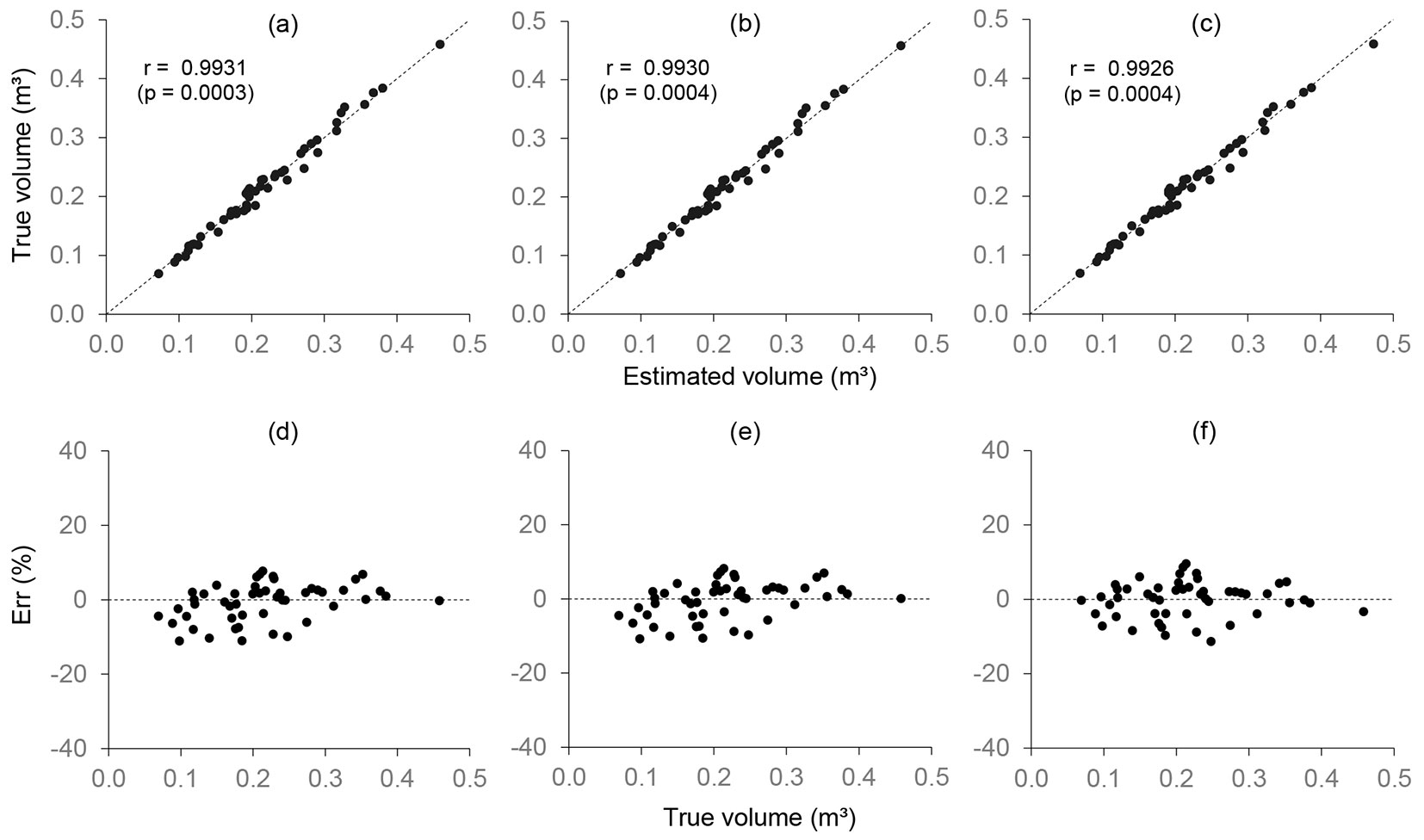
A validation of these equations revealed that the estimated volumes were not statistically different to the true volume obtained by the xylometer method (p > 0.8523) (Tab. 3). The three equations provided accurate volume estimates (Syx% < 5%), with relatively strong correlations between the true values (xylometer) and the estimates (Fig. 4a, Fig. 4b, and Fig. 4c). The AD between the true values and the estimated ones was less than 0.31% (Tab. 3), and the distribution of residuals had errors of between -11.30 and 9.58% (Fig. 4d, Fig. 4e, and Fig. 4f), with no bias.
Tab. 3 - Performance of volumetric equations in estimating Eucalyptus urophylla tree volumes in relation to a (reference) volume obtained using the xylometer method. Means followed by the same letter did not significantly differ according to a Kruskal-Wallis test (α = 0.05); (n): number of trees; (AD): aggregated difference; (Syx%): residual standard error (%).
| Treatment | n | Volume (m³) | AD (m³) |
AD (%) |
Syx% | ||
|---|---|---|---|---|---|---|---|
| Mean | Min | Max | |||||
| Xylometer | 50 | 0.2161 a | 0.0691 | 0.4581 | - | - | - |
| Xylometer equation | 50 | 0.2161 a | 0.0721 | 0.4593 | 0.0011 | 0.01 | 4.91 |
| Smalian equation | 50 | 0.2154 a | 0.0722 | 0.4577 | 0.0337 | 0.31 | 4.97 |
| (Smalian + Paraboloid) equation | 50 | 0.2160 a | 0.0692 | 0.4731 | 0.0078 | 0.07 | 4.94 |
Fig. 4 - Correlations between true (xylometer) and estimated volumes, and the distribution of residuals from volumetric equations. The following methods were used: xylometer (a and d), Smalian (b and e), and Smalian + Paraboloid (c and f).
Discussion
Rigorous scaling using the traditional Smalian’s method resulted in less variation in tree volume than that obtained by the xylometer method, so had better precision, although they were not significantly different. This outcome was expected, because the method is also used as a reference when obtaining wood volume in routine forest inventories in Brazil ([21], [19]). Smalian’s method is used in both native forests and plantations ([40]); however, it often gives less satisfactory results than other scaling methods, such as the traditional formulas of Huber and Newton ([8], [22]). Smalian’s method gives its worst estimates when combined with long and large sections, a problem that did not occur for the shorter sections ([22], [19]) used in this study.
Although the Smalian + Paraboloid method gave a higher dispersion of errors than the traditional Smalian’s scaling method, the volumes obtained were concentrated around the true mean (xylometer). Around 99% of the volumes obtained by this method had errors of between -10 and 10%. However, when considering the average volume (n = 150), this method provided similar volumes to those obtained by the Smalian and xylometer methods. It overestimated the volume by less than 1%, and is therefore considered quite accurate.
The results indicate that the part of the bole above a height of 2 m had the greatest similarity to the paraboloid form, which would naturally provide a larger volume than those obtained by the cone and neiloid forms, and explains the underestimations obtained when using these two forms to describe the tree bole. To assess the stem profile, three randomly selected trees (small, medium, and large) in the forest stand were measured to ascertain whether several geometrical solids were expressed in their forms, as described by Van Laar & Akça ([45]). We found that the paraboloid was the prototype that best adjusted to the form of the stem above a height of 2 m. Tree scaling using Smalian’s method had less variation over the dispersion of errors (Fig. 2), because the large number of sections measured along the bole allowed the capture of possible variations in the form of the stem profile.
The adjustment of the Schumacher & Hall model provided accurate equations with which to estimate tree volumes, irrespective of the tree-scaling method (Xylometer, Smalian, or Smalian + Paraboloid) used. When the resulting equations were applied to an independent dataset that was not used in the adjustment of the models (validation), they all yielded similar estimates of the true (xylometer) volume. This confirms that the method proposed (Smalian + Paraboloid) was efficient in obtaining the volume, which is a variable required for the adjustment of volumetric models.
The use of the Smalian + Paraboloid method is a promising alternative for the inventories of other forest species, such as pine and acacia. It allows us to take advantage of plot randomness in the inventory area to collect representative data that will be used in the adjustment of volumetric models capable of generating precise estimates of the volumes of other trees in the forest stand. By adopting this method, the gathering of all necessary information can be performed simultaneously with plot measurement, and diameters can be measured in a few sample trees up to a height of 2 m, thereby avoiding tree felling or the use of specialized equipment to measure standing trees.
This procedure also allows a reduction in the number of diameters measured along the bole when compared with traditional tree scaling. In this study, traditional tree scaling by Smalian’s method required the measurement of 28 diameters (on average) along the bole of each tree, while the Smalian + Paraboloid method required only 8 diameters, enabling us to reduce the number of measurements by 71%. Therefore, using this method may improve data collection and reduce the costs of forest inventory activities. In addition, it offers an alternative for obtaining tree volumes in situations where it is impractical to perform tree scaling using destructive methods.
However, some care must be taken when using this method, particularly regarding tree form, because according to Soares et al. ([41]), bole volume is strongly correlated with its form. Several factors affect tree form, including species, site, silvicultural practice, age, and genetic expression, among others ([45], [5]). Tree form in the population studied was considered similar to Apollonius’s paraboloid. However, it is important to note that the tree boles of other Eucalyptus stand, or of any other species, can resemble alternative forms of a paraboloid, or even of other geometrical solids (dendrometric prototypes). Therefore, it is important to examine each specific case, and to adopt the most appropriate method for calculating the volume. In this study, we proposed taking an average tree of the forest stand and obtaining its bole volume (vn) by applying Smalian’s method for a neiloid up to a height of 2 m. Above that height, a generic paraboloid should be used as defined in eqn. 3, i.e., v2 = g2 h2 f2. After sectioning the tree, we can obtain its volume using a xylometer in such a way as to provide the volumes for the neiloid section (vxn) and for the bole above 2 m (vx2) separately. If it is assumed that vx2 is equivalent to that of a cylinder that has a constant cross-sectional area gxc and height hx2, then we have vx2 = gxc hx2. Assuming that we wish to set the paraboloid such that the two volumes are equal (i.e., vx2 = v2), then we have hx2 = h2. Consequently, the form factor fi can be obtained experimentally, as shown in eqn. 5:
where gxc is the constant cross-sectional area of an equivalent cylinder of the bole from 2 m to the top of the tree, g2 is the cross-sectional area measured at a height of 2 m, and fi is the form factor of the bole measured above 2 m that is obtained experimentally.
Using the volumes of an average tree obtained by the xylometer method and the Smalian + Paraboloid method, as shown in Tab. 1, we calculated what was proposed above (Tab. 4).
Tab. 4 - Comparison of bole-volume estimates of an average tree taken from a Eucalyptus urophylla forest stand among different paraboloid forms. (vn): neiloid volume obtained by Smalian’s method; (v2): xylometer volume, or from Smalian, or from a different paraboloid; (vxn): neiloid volume by xylometer below 2 m; (fi): paraboloid form factor. Values followed by the same letter did not significantly differ at the 95% probability level.
| Treatment | n | Average volume (m³) | f i | ||
|---|---|---|---|---|---|
| v n | v 2 | Total | |||
| Xylometer | 150 | 0.03555 | 0.18185 | 0.2174 a | - |
| Smalian | 150 | 0.03506 | 0.18144 | 0.2165 a | - |
| Smalian + Apollonius Paraboloid | 150 | 0.03506 | 0.18397 | 0.2190 a | 0.500 |
| vxn + Apollonius Paraboloid | 150 | 0.03555 | 0.18397 | 0.2195 a | 0.500 |
| vxn+ Cubic Paraboloid | 150 | 0.03555 | 0.22025 | 0.2558 c | 0.600 |
| vxn + Semi-cubic Paraboloid | 150 | 0.03555 | 0.15735 | 0.1929 b | 0.429 |
| vxn + Equivalent Paraboloid | 150 | 0.03555 | 0.18184 | 0.2174 a | 0.494 |
From the calculation of the cross-sectional area of the cylinder, the volume of which was equivalent to that obtained above 2 m by the xylometer method, and knowing that the total height of the tree was 25.67 m (h) and the height of the paraboloid was 23.67 m (h2), then we have (eqn. 6):
From a calculation of the base cross-sectional area of the Apollonius paraboloid, we have (eqn. 7):
where g2 is the cross-sectional area measured at a height of 2 m on the stem, v2 is the volume of the bole measured above 2 m, f2 is the form factor of the bole measured above 2 m that is obtained experimentally, and h2 is the height of the tree measured above 2 m.
Therefore, the average form factor (fpe) of a paraboloid with a volume equivalent to that obtained by a xylometer is given by (eqn. 8):
where fpe is the average form factor of a paraboloid equivalent to the xylometer volume, gxc is the constant cross-sectional area of an equivalent cylinder of the bole from 2 m to the top of the tree, and g2 is the cross-sectional area measured at a height of 2 m on the bole.
As can be seen, the initial proposal of applying the Apollonius paraboloid to the data was validated. However, this approach would not always be appropriate. If a cubic or semi-cubic paraboloid was applied instead, the results would be statistically different to the parametric value, as is evident in Tab. 4.
In order to validate the methodology proposed, a sample of 28 trees was taken in another E.urophylla forest stand. After calculating the average diameter of this sample, the tree closest to this value, which in this case was the 11th tree of the sample, was identified. The data are presented in Tab. 5.
Tab. 5 - Volumes obtained for validation using the proposed methodology. (d): diameter at 2 m; (h): height; (v1): volume (m³) ≤ 2 m (Smalian); (v2): volume (m³) ≥ 2 m to the tip (Smalian); (v3): volume (m³) ≥ 2 m to the tip (Apollonius paraboloid); (tv1): total volume (m³) Smalian (traditional); (tv2): total volume (m³) paraboloid (proposed).
| Tree | d (cm) | h (m) | v1 (m³) | v2 (m³) | v3 (m³) | tv1 (m³) | tv2 (m³) |
|---|---|---|---|---|---|---|---|
| 1 | 6.70 | 14.30 | 0.00742 | 0.02152 | 0.02168 | 0.02893 | 0.02910 |
| 2 | 7.95 | 15.30 | 0.02012 | 0.01962 | 0.02261 | 0.03974 | 0.04273 |
| 3 | 7.95 | 15.90 | 0.01025 | 0.03529 | 0.03312 | 0.04554 | 0.04337 |
| 4 | 11.45 | 24.70 | 0.03052 | 0.09315 | 0.09620 | 0.12367 | 0.12671 |
| 5 | 11.75 | 21.60 | 0.02289 | 0.09792 | 0.09887 | 0.12080 | 0.12176 |
| 6 | 12.04 | 22.30 | 0.02230 | 0.10704 | 0.10743 | 0.12935 | 0.12973 |
| 7 | 12.05 | 23.50 | 0.02476 | 0.11874 | 0.11803 | 0.14350 | 0.14278 |
| 8 | 12.40 | 23.80 | 0.02421 | 0.12726 | 0.12270 | 0.15147 | 0.14691 |
| 9 | 13.05 | 24.50 | 0.02865 | 0.14182 | 0.14196 | 0.17047 | 0.17061 |
| 10 | 13.80 | 24.20 | 0.03017 | 0.14737 | 0.15030 | 0.17754 | 0.18047 |
| 11 | 14.50 | 25.50 | 0.03469 | 0.18151 | 0.18129 | 0.21620 | 0.21599 |
| 12 | 14.50 | 26.20 | 0.03469 | 0.19539 | 0.19023 | 0.23007 | 0.22491 |
| 13 | 14.53 | 24.50 | 0.03355 | 0.16289 | 0.16618 | 0.19643 | 0.19972 |
| 14 | 14.58 | 25.40 | 0.03519 | 0.17913 | 0.18025 | 0.21432 | 0.21544 |
| 15 | 14.59 | 26.10 | 0.03754 | 0.19129 | 0.19262 | 0.22883 | 0.23016 |
| 16 | 14.78 | 25.70 | 0.03715 | 0.18767 | 0.19037 | 0.22482 | 0.22752 |
| 17 | 14.88 | 29.10 | 0.03541 | 0.23071 | 0.22421 | 0.26612 | 0.25962 |
| 18 | 14.90 | 26.30 | 0.03778 | 0.20143 | 0.20015 | 0.23921 | 0.23793 |
| 19 | 14.95 | 25.60 | 0.03496 | 0.19646 | 0.19390 | 0.23142 | 0.22886 |
| 20 | 15.03 | 25.80 | 0.03565 | 0.20123 | 0.20262 | 0.23687 | 0.23827 |
| 21 | 15.30 | 26.80 | 0.03861 | 0.21367 | 0.21491 | 0.25228 | 0.25352 |
| 22 | 16.12 | 29.30 | 0.04203 | 0.25962 | 0.25992 | 0.30164 | 0.30194 |
| 23 | 16.40 | 29.20 | 0.04468 | 0.26931 | 0.26953 | 0.31399 | 0.31421 |
| 24 | 16.78 | 27.10 | 0.04532 | 0.26122 | 0.26405 | 0.30654 | 0.30937 |
| 25 | 17.57 | 29.70 | 0.05032 | 0.31655 | 0.32063 | 0.36686 | 0.37094 |
| 26 | 19.20 | 30.00 | 0.06251 | 0.38638 | 0.39140 | 0.44889 | 0.45991 |
| 27 | 19.50 | 29.40 | 0.06823 | 0.37175 | 0.37701 | 0.43998 | 0.44924 |
| 28 | 21.23 | 32.90 | 0.07183 | 0.51651 | 0.52842 | 0.58835 | 0.60025 |
The cross-sectional area of the cylinder, the volume of which is equivalent to the volume of the paraboloid obtained by Smalian’s method, is given by (eqn. 9):
where gxc is the constant cross-sectional area of an equivalent cylinder of the bole from 2 m to the top of the tree, v2s is the volume of the bole measured above 2 m equivalent to the volume of the paraboloid obtained by Smalian’s method, and ht is the total height of the tree.
The cross-sectional area of the base of the paraboloid is obtained by (eqn. 10):
where g2 is the cross-sectional area measured at a height of 2 m on the stem and d2 is the diameter measured at 2 m.
The form factor of the paraboloid to be used for the calculation of the volumes of the remaining sample trees is obtained by (eqn. 11):
where fi is the form factor of the paraboloid to be used to calculate the volumes of the remaining trees, gxc is the constant cross-sectional area of an equivalent cylinder of the bole from 2 m to the top of the tree, and g2 is the cross-sectional area measured at a height of 2 m on the bole.
As can be seen, the form factor for this new sample resulted in a value that was even closer to 0.5, i.e., the Apollonius paraboloid was appropriate for the calculation of the volumes of the remaining trees. The validation results are presented in Tab. 5.
The distribution of residuals for the volume obtained by Smalian’s method had errors of between -7.53 and 4.75%, with no evidence of bias. The correlation coefficient between the volumes obtained by Smalian’s method and the proposed paraboloid was 0.9997; see Fig. 5.
Fig. 5 - Correlations between volumes obtained by Smalian’s method and the proposed paraboloid method (a). Distribution of residuals (b). Histogram of residuals (c).
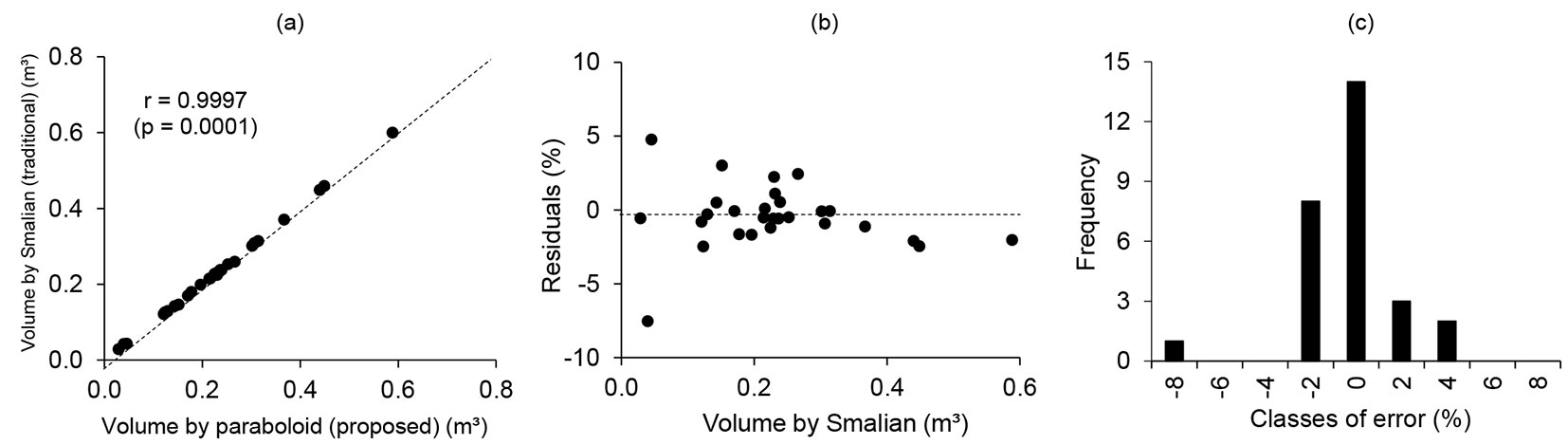
The analytical solution obtained by using a paraboloid in the second section of the tree above 2 m supports our first hypothesis. The proposed methodology for finding an appropriate form factor of a paraboloid to be used in the calculation of the volume equivalent to that obtained by a xylometer was efficient and accurate, which supports our second hypothesis.
Conclusions
The values obtained by traditional scaling using Smalian’s method (destructive) and the Smalian + Paraboloid method (non-destructive) were not statistically different to those obtained by the reference method (xylometer); therefore, they are equivalent.
The adjustment of the Schumacher & Hall model with data obtained by compatible methods (Xylometer, Smalian, and Smalian + Paraboloid) enabled us to obtain accurate equations for estimating tree volume in a E. urophylla stand.
The Smalian + Paraboloid method (without tree felling) would improve data collection and reduce the cost of the forest inventory activities that are conducted in forest plantations, and the method can obtain tree volumes in situations in which scaling (destructive) is impracticable.
Updating the volumetric estimators for older E. urophylla forest stands would be simple and inexpensive, and of major importance in ensuring the accuracy of volumetric estimators for old trees.
The use of an equivalent paraboloid to express the mean volume of trees sampled by the xylometer or traditional Smalian’s method maximizes the accuracy of volume estimation in stands of E. urophylla.
The methodology proposed for E. urophylla is suitable for other species, given the high uniformity and homogeneity of forest plantations in Brazil.
Acknowledgments
The authors would like to thank the Mixed Cooperative of Araguaia Valley (COMIGO) for financial support, providing the study site, and support during the field measurements.
EPM designed and setup the study. EPM, GBA, GTOSA, and SPN analyzed the data and wrote the paper. AVR and RSP reviewed the paper.
References
CrossRef | Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Alba Valéria Rezende
Reginaldo Sérgio Pereira
Department of Forest Science, University of Brasília (UnB), Campus Darcy Ribeiro, Brasília, CEP 70910-900 (Brazil)
Department of Forest Science, Federal University of Paraná (UFPR), Av. Prof. Lothário Meissner, 900 Campus III UFPR - Botanical Garden, Curitiba, CEP 80210-170 (Brazil)
Glauce Taís de Oliveira Sousa Azevedo
Federal University of Mato Grosso do Sul (UFMS), Campus Chapadão do Sul, Rod. MS 306, Km 105, CXP 112, Chapadão do Sul/MS, CEP 79560-000 (Brazil)
Corresponding author
Paper Info
Citation
Miguel EP, Péllico Netto S, Azevedo GB, Azevedo GTOS, Rezende AV, Pereira RS (2018). Alternative methods of scaling Eucalyptus urophylla trees in forest stands: compatibility and accuracy of volume equations. iForest 11: 275-283. - doi: 10.3832/ifor2155-011
Academic Editor
Davide Travaglini
Paper history
Received: Jun 27, 2016
Accepted: Jan 09, 2018
First online: Mar 29, 2018
Publication Date: Apr 30, 2018
Publication Time: 2.63 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2018
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 50180
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 41731
Abstract Page Views: 3322
PDF Downloads: 4142
Citation/Reference Downloads: 18
XML Downloads: 967
Web Metrics
Days since publication: 2880
Overall contacts: 50180
Avg. contacts per week: 121.97
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2018): 5
Average cites per year: 0.63
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Review Papers
Integration of forest mapping and inventory to support forest management
vol. 3, pp. 59-64 (online: 17 May 2010)
Research Articles
Simplified methods to inventory the current annual increment of forest standing volume
vol. 5, pp. 276-282 (online: 17 December 2012)
Research Articles
Tree volume modeling for forest types in the Atlantic Forest: generic and specific models
vol. 13, pp. 417-425 (online: 16 September 2020)
Research Articles
Allometric relationships for predicting the stem volume in a Dalbergia sissoo Roxb. plantation in Bangladesh
vol. 3, pp. 153-158 (online: 15 November 2010)
Commentaries & Perspectives
Benefits of a strategic national forest inventory to science and society: the USDA Forest Service Forest Inventory and Analysis program
vol. 1, pp. 81-85 (online: 28 February 2008)
Research Articles
Estimation of aboveground forest biomass in Galicia (NW Spain) by the combined use of LiDAR, LANDSAT ETM+ and National Forest Inventory data
vol. 10, pp. 590-596 (online: 15 May 2017)
Research Articles
Integrating area-based and individual tree detection approaches for estimating tree volume in plantation inventory using aerial image and airborne laser scanning data
vol. 10, pp. 296-302 (online: 15 December 2016)
Research Articles
Comparing land use registry and sample based inventory to estimate forest area in Podlaskie, Poland
vol. 10, pp. 315-321 (online: 23 February 2017)
Technical Advances
Improved estimates of per-plot basal area from angle count inventories
vol. 7, pp. 178-185 (online: 17 February 2014)
Research Articles
Using self-organizing maps in the visualization and analysis of forest inventory
vol. 5, pp. 216-223 (online: 02 October 2012)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







