
Density and spatial distribution of beech (Fagus sylvatica L.) regeneration in Norway spruce (Picea abies (L.) Karsten) stands in the central part of the Czech Republic
iForest - Biogeosciences and Forestry, Volume 9, Issue 4, Pages 666-672 (2016)
doi: https://doi.org/10.3832/ifor1581-008
Published: Mar 12, 2016 - Copyright © 2016 SISEF
Research Articles
Abstract
The conversion of Norway spruce monocultures to mixed forest is a long-term and costly process, and little information is available on the silvicultural use of spontaneous regeneration of native tree species to such purposes. In this context, we focused on the natural expansion of European beech in pure Norway spruce stands currently occurring in central Europe. The study was conducted in three secondary spruce stands with single adult (seed) beech trees growing on acidic sites of the fir-beech vegetation zone in the Bohemian-Moravian Highlands (Czech Republic). The regeneration strategy of beech and spruce (in terms of density and height increment) was studied under different light regimes (expressed using the indirect site factor - ISF) on the forest edge (9.7-41.2% ISF) and within gaps in the forest interior (10.7-20.8% ISF). Beech showed a broad light adaptability, being present under the spruce canopy with high density, depending on the distance from beech seed trees. Sparse beech saplings were found at a distance of more than 100 m from adult beech trees; however, abundant beech regeneration was found up to a distance of 25 m. Contrastingly, the density and growth in height of the spruce regeneration was strongly affected by light conditions. Spruce reached a density higher than beech in the understory where ISF > 17% (i.e., up to a distance of 45 m from the stand edge or within gaps no smaller than 400 m2), and competed with beech in terms of height growth where ISF > 20% (i.e., at the very stand edges). Knowledge of the spatial pattern and the light strategy of both species provided useful information to support the conversion of spruce monocultural stands to mixed spruce-beech forests.
Keywords
Natural Regeneration, Beech, Spruce Monoculture, Competition, Conversion, Mixed Forest
Introduction
Utilization of natural processes in forest management is one of the major challenges for ecologically-based forestry today. The central European model of cultivating broadleaves for converting unstable, often pure coniferous stands, to close-to-nature mixed forests is known and relatively well-documented ([42], [10]). Few data exist, however, on the use of forest succession elements for management purposes. Remnants of adult individuals and groups of native species, primarily broadleaves (e.g., oak, beech and maple) and/or conifers (such as fir), admixed in pure secondary coniferous stands, can be found both in Europe ([20], [11], [24], [45], [7], [8]) and in other parts of the world ([15], [16]). Their potential for spontaneous reproduction differs depending on climatic, site and stand conditions, as well as the silvicultural strategy. While Küßner ([25]) in the eastern part of the Ore Mts. (Germany) and Diaci ([5]) in the Slovenian Alps reported a slow succession of climax woody plants in spruce plantations due to the low number of fruiting individuals within populations, Dobrovolný & Tesar ([7], [8]) recorded an expansion of beech under the shelter of spruce stands in the Czech Republic and in the central Saxon part of the Ore Mts. (Germany). Today, in central Europe the percentage of beech is increasing spontaneously to the detriment of conifers both in managed forests ([44]) and in old-growth ([21]).
The spontaneous penetration of oak into pine stands due to a process referred to as “jay (Garrulus glandarius) seeding” is well documented ([31], [45]). Oak saplings can be found up to several kilometers from the seed tree ([3], [22], [12]). Similarly, spontaneous spreading of beech occurs in spruce stands of central Europe, though it has not been sufficiently documented yet. Beech seeds can be transported to a distance of several dozen meters by rodents ([18]) and even to longer distances by jays ([19], [24], [49]). Although the dissemination distance may exceed 100 m ([11], [17]) or even several kilometers ([19], [24]), the largest amount of beech seeds is frequently found up to 20 m from the seed tree ([48], [26], [39], [49]).
The successful expansion of beech in the past 20-30 years has been partly attributed to more frequent and abundant masting, occurring at 2 to 5-year intervals ([14], [32], [40]). During mast years, the abundance of beech seeds can be as high as 300-1000 seeds m-2 ([38]). Even old or suppressed beech trees produce seeds in such periods ([2], [37], [7]).
Besides seed production and ecology, natural regeneration of beech and its competitive ability are influenced by many biotic and abiotic factors such as animals, mildew, stand conditions, climatic factors, soil moisture and supply of nutrients, root competition, ground vegetation, etc. ([30], [1]). Nevertheless, light obviously plays the key role in the natural regeneration of this species ([33], [49]). Shade tolerance gives beech a competitive advantage over other species ([33], [30], [1], [49]). Indeed, beech can survive under extremely low radiation (3-5%) for several years, with slow growth rate but a very fast response to release ([4], [49]). Height and diameter increments are highest at relative light intensity (RLI) of 100%, with only a slight decrease occurring at 30%<RLI<50% ([49]). In general, the relative radiation should not fall below 5-10 % in order to maintain beech height growth and stem quality ([13], [28], [6]). It has been reported that beech maintains its advantage over spruce when the relative diffuse radiation (RDR) is below 15% (corresponding approximately to a stand basal area of 30-35 m2), while at values higher than 20%, spruce becomes dominant in terms of height growth ([27], [48], [29], [23], [43]).
The main goal of this study was to better understand the natural regeneration process of beech as compared with spruce in a Norway spruce monoculture which is being converted to a mixed forest. Specific goals were (i) to quantify the occurrence and density of beech regeneration in relation to the distance from seed trees; and (ii) to determine the dependence of the occurrence and height growth of beech and spruce regeneration on light conditions. Finally, the results should be used for silvicultural recommendations.
Material and methods
The research was carried out in the Bohemian-Moravian Highlands, the largest region of beech and fir-beech vegetation of the central part of Czech Republic. The region is currently covered by spruce plantations (about 70 % of the forest area) growing on acidic soil (Cambisols) at middle elevations, with an average total precipitation of 700 mm and a mean annual temperature of about 6 oC.
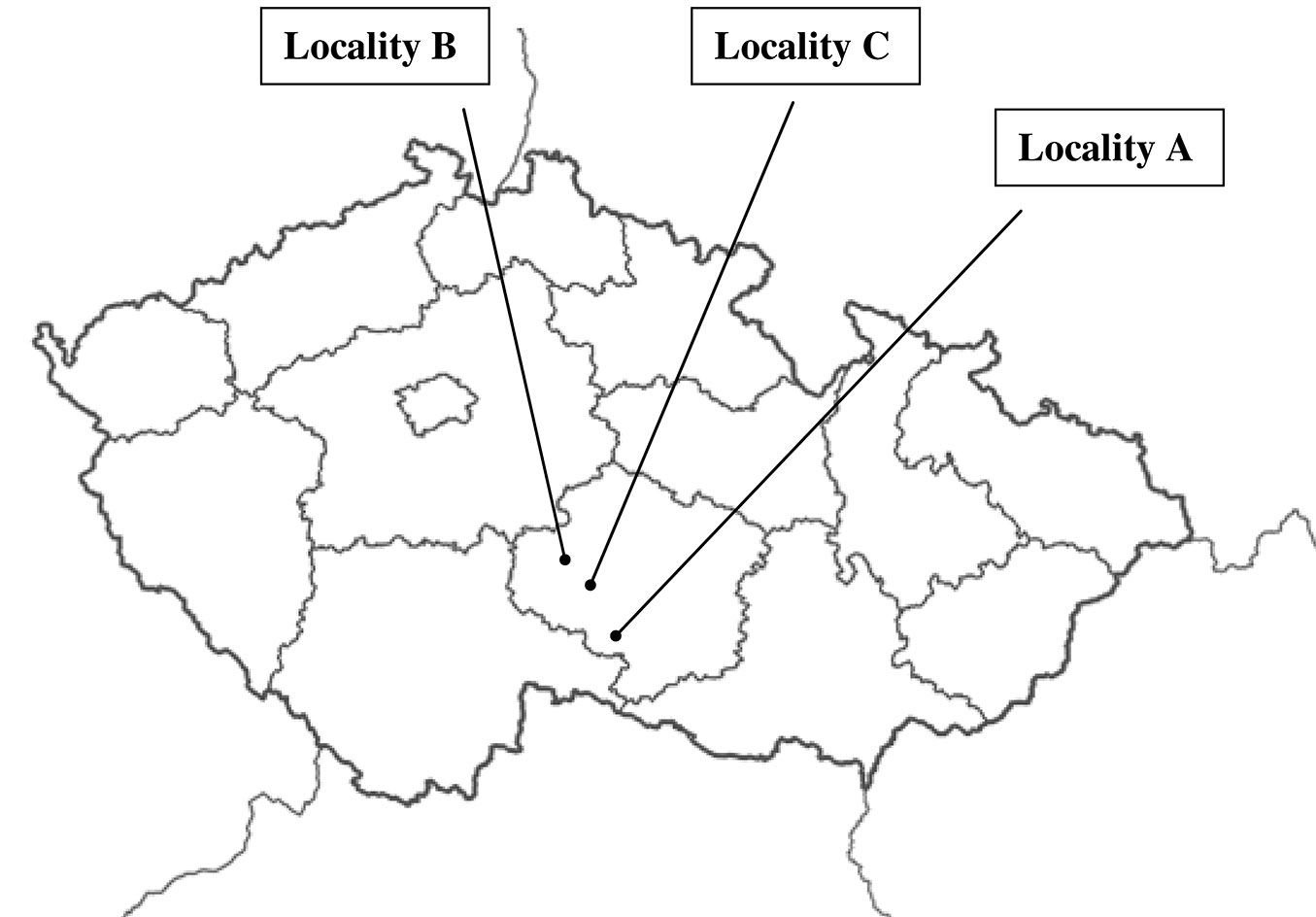
Three managed forest stands (A, B, C) characterized by adult spruce monocultures with irregular admixture of single adult beech trees were selected in the study area (Tab. 1, Fig. 1). The stocking density was around 80 % of the standard volume, due to salvage felling as a result of occasional damage by icing and bark beetles.
Tab. 1 - Characheristics of the studied stands.
| RP | Stand | Floristic association/ type of site |
Altitude (m a.s.l.) |
Area (ha) | Age (years) | Volume (m3 ha-1) |
Species composition | Location |
|---|---|---|---|---|---|---|---|---|
| A | 957F11 | Acidic beech-silver fir forest | 560 | 6.60 | 109 | 577 | spruce 99%, oak 1% | 49°17′ 34″ N 15°42′ 53″ E |
| B | 829E 12 | Acidic beech-silver fir forest | 640 | 6.49 | 112 | 672 | spruce 93%, beech 4%, Scots pine 1%, larch 1%, Douglas-fir 1% |
49°23′ 14″ N 15°42′ 43″ E |
| C | 33a10 | Acidic beech-silver fir forest | 640 | 5.52 | 102 | 563 | spruce 95%, Douglas-fir 5% | 49°24′ 47″ N 15°19′ 16″ E |
Fig. 1 - Location of the three study areas.
For each stand, the spatial pattern of distribution of beech regeneration up to 2 m in height was recorded, as well as the density and the height growth of seedlings and saplings of beech and spruce in different light environments (Fig. 2). Measurements were conducted in the summer of 2013 along two transects, the first staked between two adult beech trees (A: 52 m; B: 72 m; C; 54 m), while the second was starting at an adult beech tree and directed toward the homogeneous, pure spruce stand with no adult beech trees, up to a distance of 150 m (Fig. 2). Both transects were divided into sections of 2 m in length and the density of beech regeneration was determined in circular 1-m2 subplots centered in the middle of each section. The total number of subplots was 101 at the site A, 111 at the site B, and 102 at the site C.
Fig. 2 - Graphical representation of the experimental design adopted in this study.
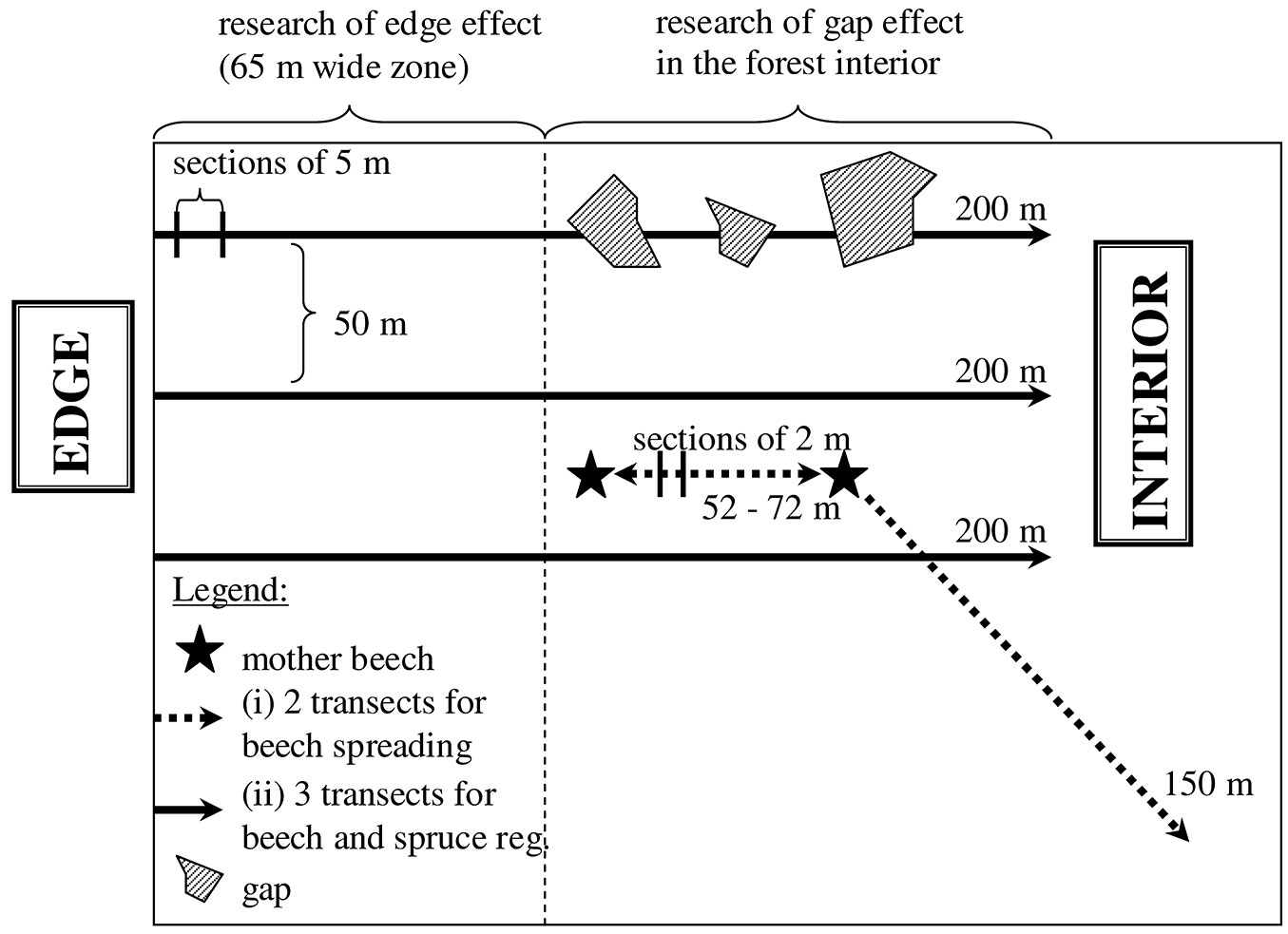
Measurements were conducted in two distinct areas with different light conditions: (i) in the 65 m-wide outermost zone of the forest (EDGE); (ii) in the inner part of the stands (INTERIOR). To this purpose, measurements were carried out along three transects set at a distance of 50 m each other (Fig. 2). The transects were staked perpendicular to the forest edge and directed towards the interior of the stand, their total length being 200 m (variant EDGE up to 65 m and variant INSIDE the next 135 m). Each variant was divided into sections of 5 m in length and a circular 5-m2 subplot was established in the middle of each section. The total number of subplots established in each stand was 39 for the variant EDGE and 81 for the variant INSIDE. For each subplot, the following attributes of the regeneration of beach and spruce were measured: (i) density (n ha-1); (ii) height of the apical bud above the ground (in cm); (iii) annual height increment of the leading shoot from the previous year (in cm). In the variant INTERIOR, the spatial positions of the canopy gaps above these transects were measured using the Field-Map technology (Institute of Forest Ecosystem Research, Ltd., Czech Republic). Canopy gaps were classified in the following classes: 25, 50, 100, 400 m2. Hemispherical photographs were also taken in the middle of each section (of both variants) using a digital Nikon Coolpix 4500® camera with the FC-E8 fisheye converter. The indirect site factor ISF (i.e., the intensity of relative diffuse radiation) was evaluated using the software WinsCanopy® 2008a (Regent Instruments Inc., Canada). In the case of variant EDGE, the ISF values ranged from 9.7 to 41.2%, while for the variant INTERIOR ISF ranged from 10.7 to 20.8 %.
Statistical analyses were carried out using the software package STATISTICA® ver. 10 (StatSoft Inc., Tulsa, OK, USA). The Kruskal-Wallis one-way analysis of variance was used to test for differences in the regeneration variables between sites and variants. Binary logistic regression procedures were used to verify the relationship between selected factors (i.e., distance from the adult beech tree, ISF and distance from the edge) and the occurrence of beech and spruce regeneration. Binary logistic regression estimates the probability (0-1) of occurrence of a selected characteristic (e.g., the probability of “success”). For this analysis, the presence of saplings was used as a dependent variable. All the plots were classified according to the occurrence of beech or spruce regeneration, where 0 means no regeneration and 1 means one or more saplings per m2. where. The general form of the model function used in this analysis was as follows (eqn. 1):
where β0 is the intercept and β1 xi is the regression coefficient multiplied by the predictor.
Results
In general, the average density of beech regeneration in the forest interior was greater than that of spruce, though significant differences were detected only for the variant “AINTERIOR” (Tab. 2). As expected, there were significantly more favorable light conditions for spruce regeneration on the forest edge, and its density was significantly greater than for beech in all the studied stands. Beech regeneration showed higher values of height growth as compared to spruce in both variants EDGE and INTERIOR: significant differences were found among all variants except “CEDGE”. The forest edge was characterized by the highest and the more heterogeneous values of radiation compared with the forest interior. Obviously, the amount of light near the forest edge was varying based on the distance from the edge, while in the forest interior, it was affected by the sizes of the gaps (Tab. 2).
Tab. 2 - Statistics of beech and spruce regeneration at the study sites. Mean ± 95% interval values are reported according to the stand studied (A, B, C) and the variant analyzed (EDGE and INTERIOR).
| Stand | ISF (%) |
Density of Beech (N ha-1) |
Density of Spruce (N ha-1) |
Height of Beech (cm) |
Height of Spruce (cm) |
Height increment of Beech (cm) |
Height increment of Spruce (cm) |
|---|---|---|---|---|---|---|---|
| AINTERIOR | 13.4 | 35881 | 1973 | 36.9 | 24.6 | 9.6 | 2.81 |
| ±95% | 0.1 | 6303 | 919 | 3.9 | 6.5 | 1.6 | 1.6 |
| AEDGE | 16.4 | 10800 | 18263 | 64.0 | 34.8 | 15.9 | 6.0 |
| ±95% | 0.3 | 3098 | 3409 | 10.5 | 3.2 | 4.9 | 0.5 |
| BINTERIOR | 19.7 | 50556 | 126111 | 31.4 | 9.0 | 5.4 | 1.9 |
| ±95% | 0.2 | 40586 | 40901 | 7.1 | 1.6 | 1.6 | 0.3 |
| BEDGE | 24.2 | 42222 | 141944 | 50.1 | 18.0 | 9.5 | 4.2 |
| ±95% | 2.3 | 21900 | 40277 | 12.0 | 3.8 | 2.0 | 0.8 |
| CINTERIOR | 15.9 | 16200 | 8700 | 60.4 | 27.8 | 15.3 | 2.8 |
| ±95% | 0.2 | 4980 | 5729 | 5.6 | 2.9 | 1.7 | 0.8 |
| CEDGE | 25.9 | 9167 | 65556 | 39.3 | 23.0 | 7.5 | 3.8 |
| ±95% | 1.3 | 4154 | 19182 | 8.7 | 8.4 | 2.0 | 1.0 |
| AllINTERIOR | 14.6 | 32254 | 14924 | 40.9 | 19.4 | 11.3 | 2.3 |
| ±95% | 0.2 | 5659 | 5204 | 3.3 | 3.4 | 1.2 | 0.4 |
| AllEDGE | 18.0 | 17035 | 34809 | 55.3 | 31.6 | 12.5 | 5.5 |
| ±95% | 0.5 | 5211 | 6279 | 6.8 | 2.8 | 2.7 | 0.4 |
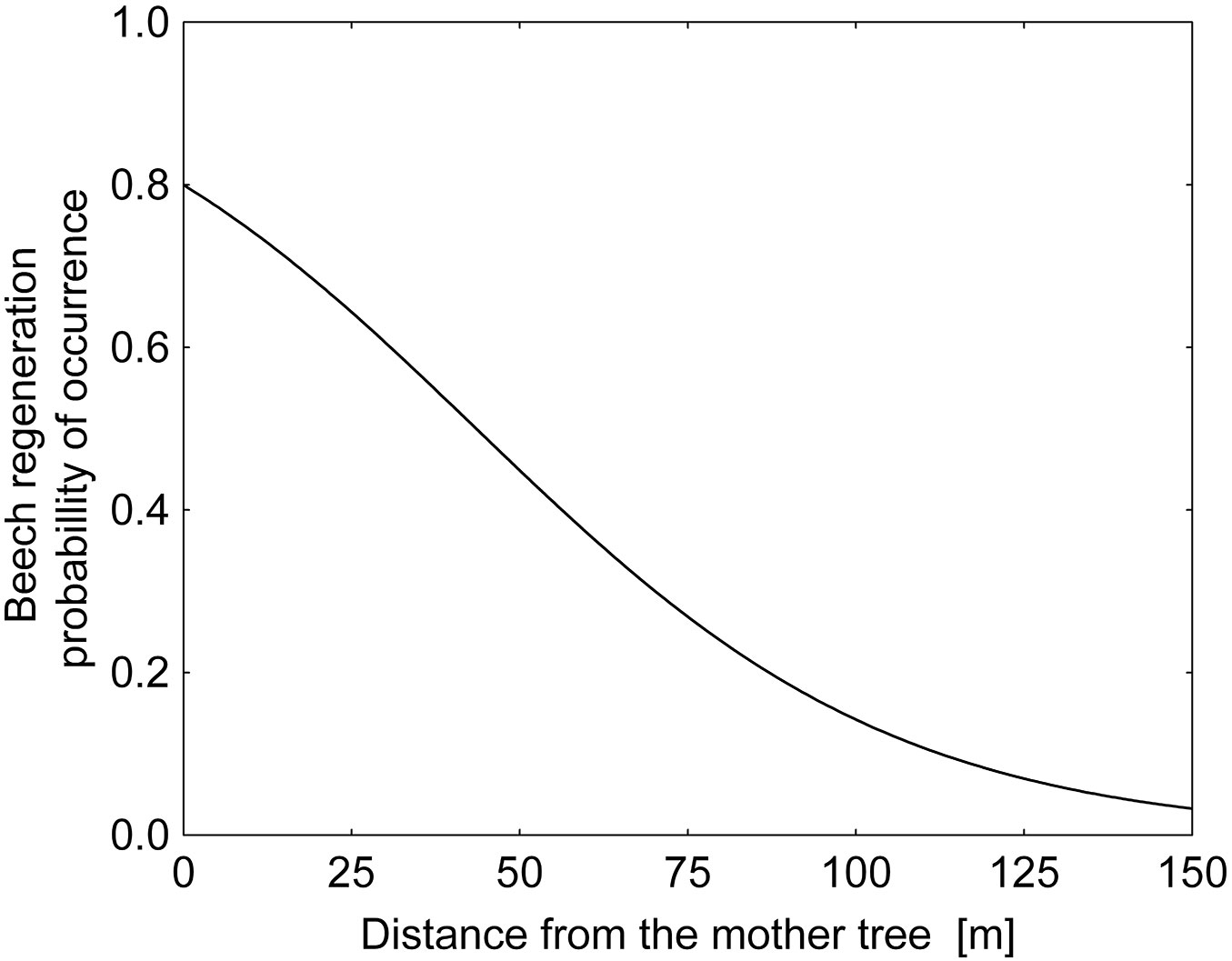
The percentage and average density of beech regeneration showed a steeply decreasing trend up to 25 m from the seed tree in all the three stands investigated (Fig. 3). The highest density of beech saplings was found directly below the crown of the seed trees (within a radius of 0.1-5 m), while the highest percentage of all saplings was recorded in the surrounding area, at a distance of 5-10 m. At larger distances (>50 m) from seed trees the sapling density was low, though single individuals were found at distances exceeding 100 m. Pooling the data from all the three stands, we constructed a general model using the significant parameters for the abundance of beech saplings as a function of distance from seed trees (Fig. 4, Tab. 3). The curve confirmed a low probability of occurrence of saplings at distances exceeding 50 m from the seed tree.
Fig. 3 - Average density of beech regeneration and percentage of the total number of individuals according to the distance from the seed tree. The highest density of regeneration was found at a distance of 0.1-5 m from seed trees, while the highest percentage of all individuals was recorded at a distance of 5-10 m.
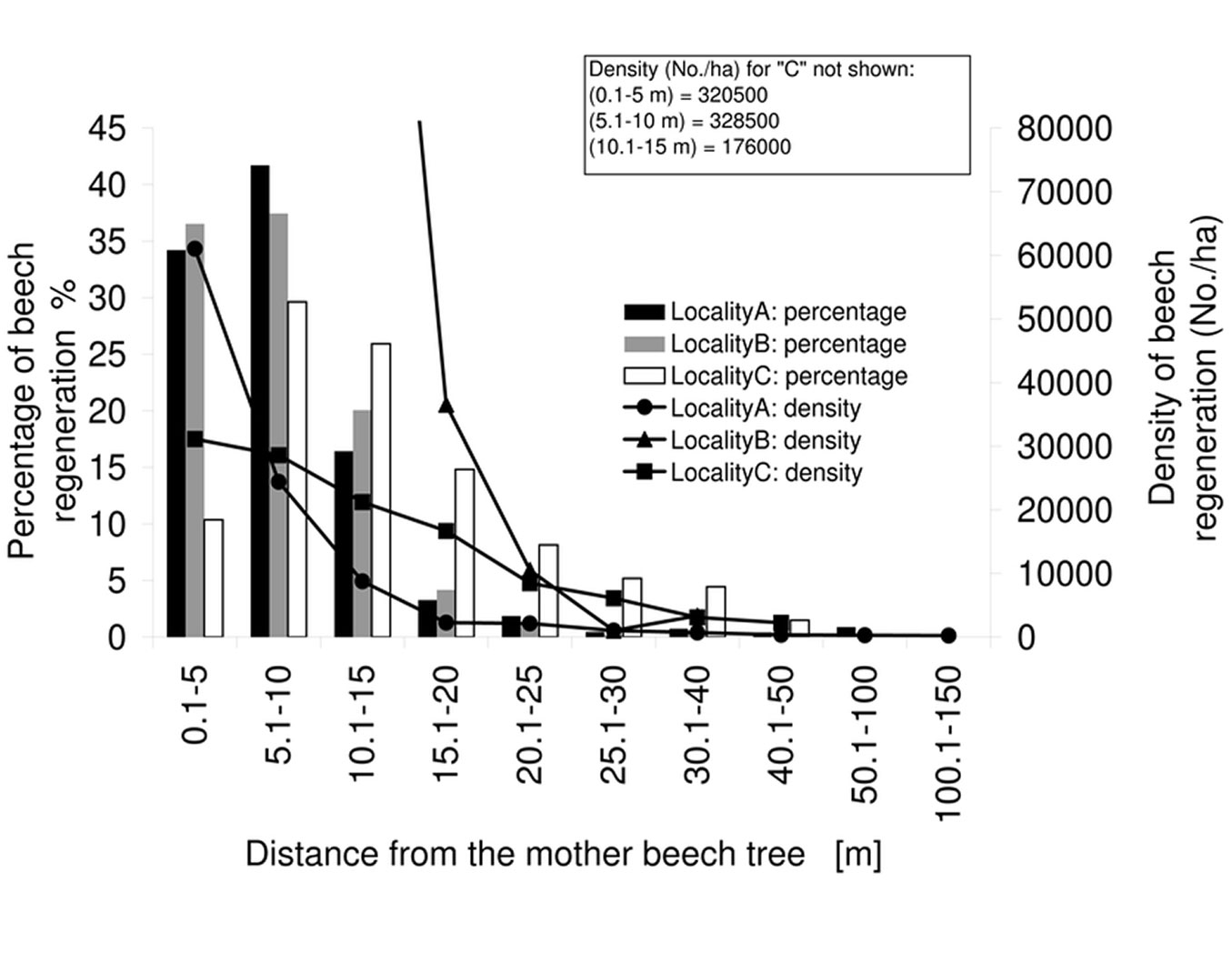
Fig. 4 - Logistic regression of beech saplings distribution. The probability of the occurrence of beech regeneration (present = 1, no regeneration = 0) as a function of the distance from the seed tree. At distances exceeding 50 m, the density of beech saplings is low and the probability of occurrence of beech regeneration falls below 0.5.
Tab. 3 - Statistics of the logistic regression.
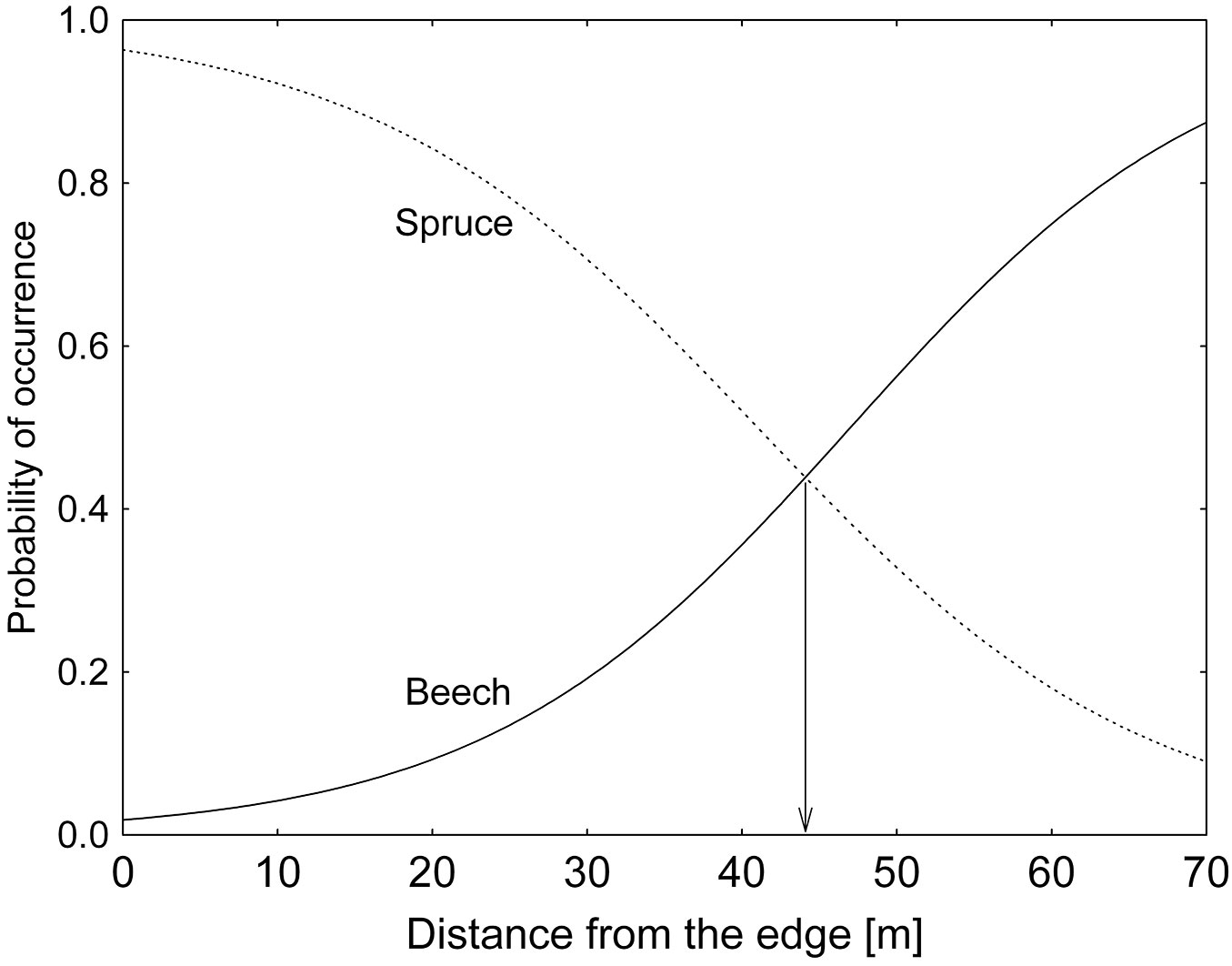
Using the logistic regression on the data pooled over the three studied stands, we calculated the approximate value of ISF (value of diffuse radiation) at which beech and spruce showed similar regeneration abundance, i.e., the competitive point for spruce and beech in terms of ISF (Fig. 5). Beech regeneration was more frequent below 17% ISF, while spruce exhibited a competitive advantage over beech above such value. As a consequence, spruce regeneration overtakes beech regeneration up to 45 m from the forest edge, as displayed in Fig. 6.
Fig. 5 - Probability of occurrence of beech and spruce regeneration (present = 1, no regeneration = 0) as a function of the diffuse radiation (ISF). As compared with spruce, the probability of occurrence of the beech regeneration resulted higher at lower diffusion radiation values in all the plots. The threshold of competitive advantage between the two species was about 17% ISF.
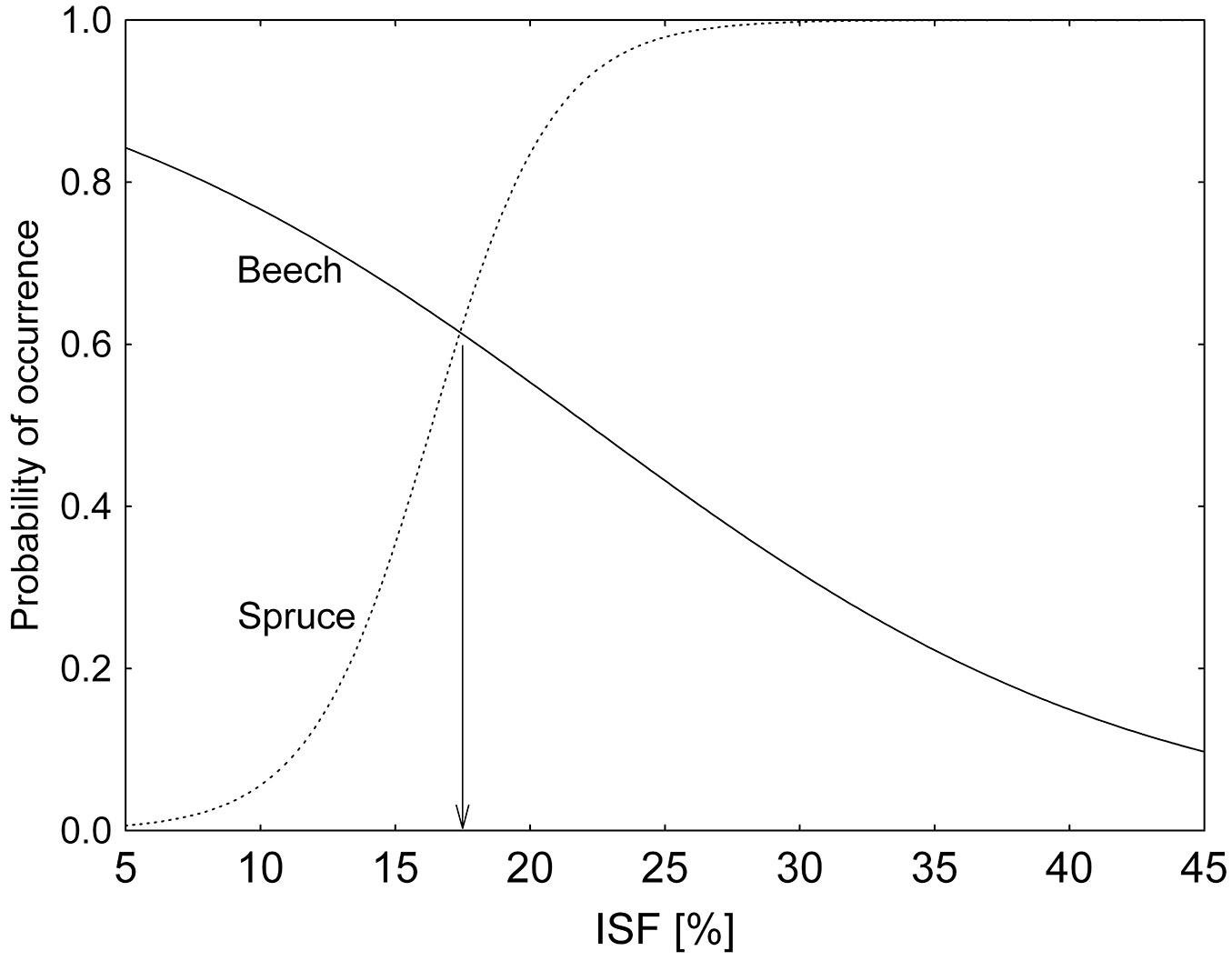
Fig. 6 - Probability of occurrence of beech and spruce regeneration (present = 1, no regeneration = 0) as a function of distance from the forest edge. The logistic regression model applied on the pooled dataset revealed that the probability of occurrence of spruce regeneration is higher than that of beech up to 45 m from the edge.
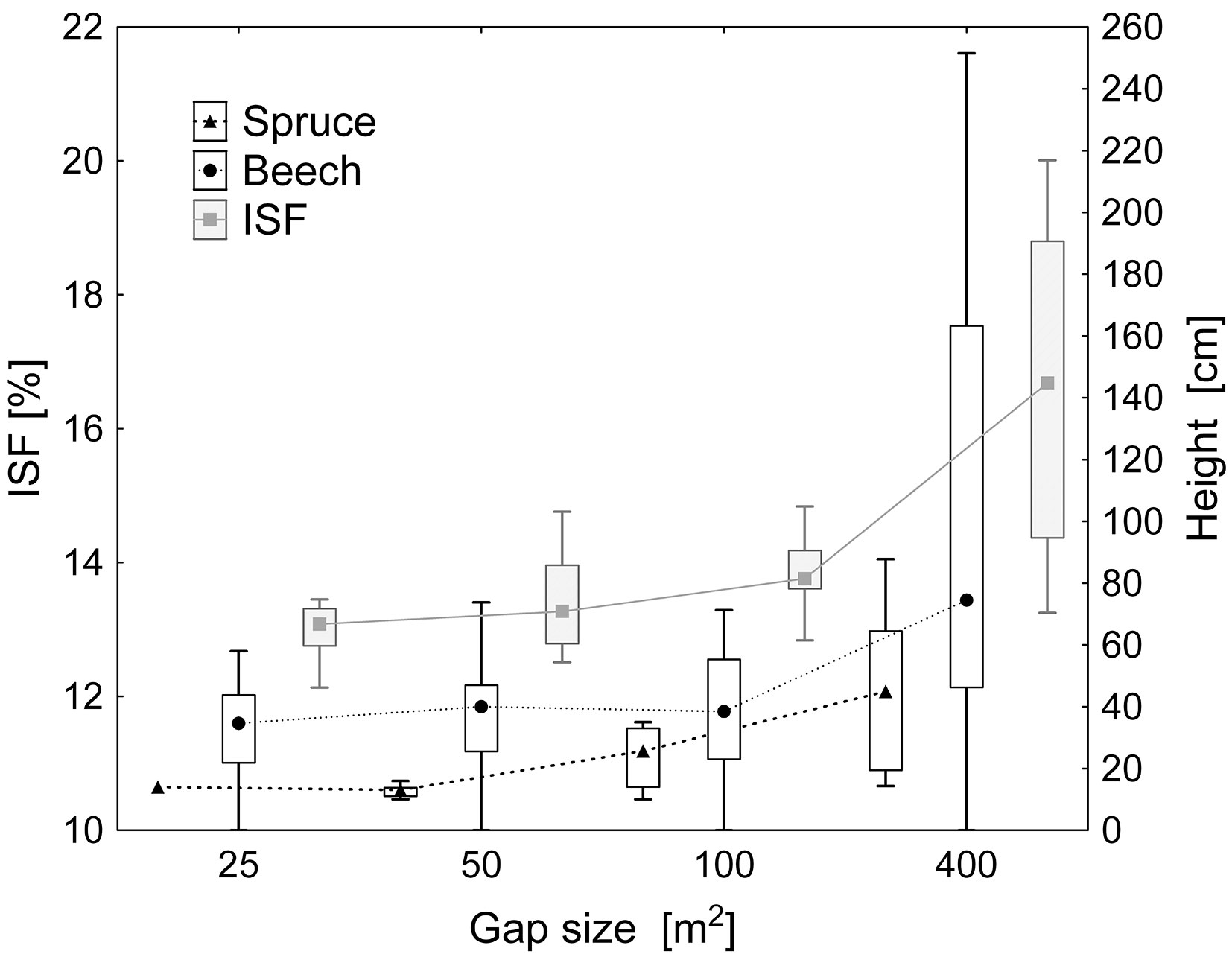
In terms of height growth, spruce competed with beech only at the very edges of the forest with ISF above 20%, otherwise beech dominated (Fig. 7). As for the forest interior, spruce was outperformed by beech in the whole range of the gap sizes (Fig. 8). Spruce regeneration started to grow only in the largest gaps of approximately 400 m2.
Fig. 7 - Diffusion radiation (ISF %) and height growth of beech and spruce regeneration at different distances from the forest edge. In terms of growth, beech outperformed spruce over the whole range of distances. Spruce showed a competitive advantage only at the very edges of the forest (ISF > 20%).
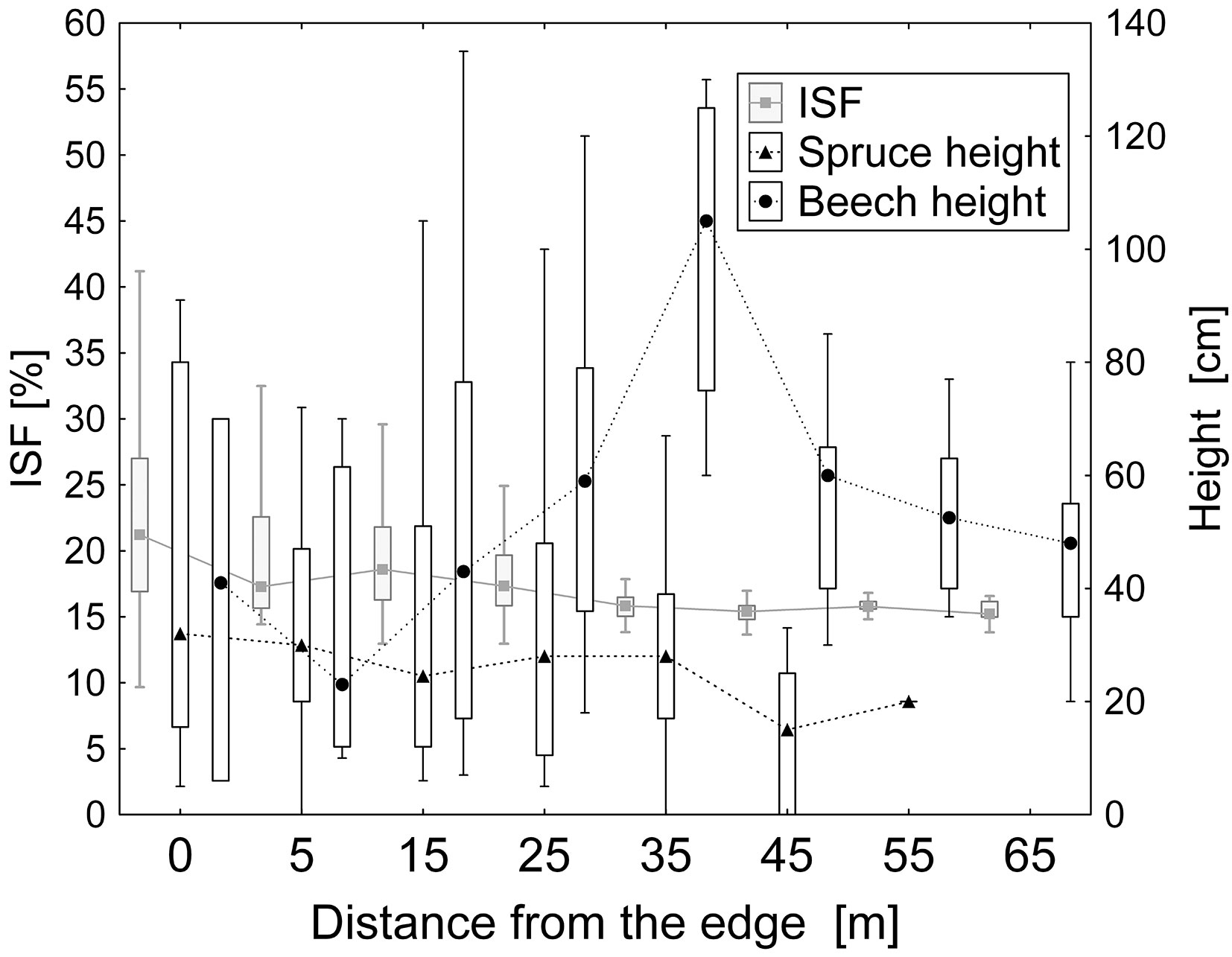
Fig. 8 - Diffusion radiation (ISF %) and height growth of beech and spruce regeneration according to the size of gaps. In terms of height growth, beech outperformed spruce over the whole range of the gap sizes. Spruce showed a significant height growth only in the largest gaps of 400 m2.
Discussion
Natural regeneration is a complex process affected by many biotic and abiotic factors. It was observed that in spruce monocultures with occasional presence of single adult beech trees, abundant regeneration of beech is expected to occur up to a distance of approx. 25 m from seed trees (Fig. 3). Similar distances from seed trees were also reported by other studies ([20], [26], [11], [49], [48]). Beech regeneration, however, may occasionally occur at much greater distances (up to 100 m and possibly farther) from the seed tree (Fig. 3, Fig. 4). Indeed, Irmscher ([17]) reported beech regeneration as far as 254 m from seed trees. All these observations clearly demonstrate the key role of animals in the dispersion of beech seeds, as already described by other studies ([47], [24], [11], [26]). From the silvicultural point of view, Dobrovolný & Tesar ([8]) consider the threshold of 20 m as the maximum distance between seed trees in order to achieve an adequate density of beech saplings (i.e., a minimum of 10 000 saplings ha-1), which is also the standard prescribed by the Czech thinning guideline ([41]). Thus, approximately 2-3 seed trees ha-1 in a spruce stand may provide for up to 30% share of beech regeneration in the subsequent stand generation. Nevertheless, the minimum management objective can be achieved even with a lower density of beech saplings, from which new seed trees will originate.
Furthermore, our research confirmed a different regeneration strategy of beech and spruce. Beech saplings with their broad light adaptability occurred in various light conditions and their density was influenced mainly by their distance from seed trees. The low correlation between light conditions and the density of beech saplings in the initial regeneration phase are pointed out in numerous works ([30], [48], [46], [23], [34], [9], [35], [36], [1]); however, the correlation was stronger with increasing age ([46]). Unlike beech, spruce requires more than 17% ISF (i.e., a 45m-wide edge zone or gaps of at least 400 m2 in the forest interior) to colonize the higher density spots and more than 20% of ISF (i.e., at the very edges) to compete with beech in height growth. The competitive threshold of approximately 20% of the diffuse radiation is confirmed by many other authors ([27], [23]). Lüpke & Spellmann ([29]) observed a competitive advantage of beech over spruce with ISF 12.7 % (10-20%) and the exact opposite with 30.1 % (20-40%). Spruce found suitable conditions only inside the gaps (15-25 m in diameter) or in more opened parts, where the basal area decreased to approx. 60%. In an adult mixed stand Kühne & Bartsch ([23]) observed only beech regeneration with a PAR site factor of 12.5% and only spruce regeneration with a mean PAR site factor of 21.2%. Within the range 0.2-28.9% of ISF, Unkrig ([48]) reports pure beech regeneration where ISF < 12 % (mean = 7.1%), pure spruce regeneration between 8.6 and 19% (mean = 15.1%) and a mixture of the two species with ISF between 4 and 20% (mean = 10.9 %).
From a practical point of view, we found that beech has no problems in regenerating after low intensity felling as compared with spruce. This is in agreement with the evidence reported by Kühne & Bartsch ([23]) on the natural regeneration of mixed stands in the submontane vegetation zone.
Conclusions
Under the natural and stand conditions explored in this study (Czech Republic - fir-beech vegetation zone - acidic site), beech showed a broad light adaptability and successfully colonized the space under the shelter of spruce canopy, depending upon the locations of adult beech trees. Despite the long distance of beech saplings from single seed trees (more than 100 m), abundant regeneration is expected to occur to a distance of 25 m. Thus, approximately 2-3 seed trees ha-1 may provide for an up to 30% share of beech saplings in the subsequent stand generation. As compared with beech, spruce showed a higher sensitivity to light, showing a higher probability of occurrence at the edge of the forest (in the 45 m-wide zone) or in the largest gaps (400 m2) of the forest interior. In terms of height growth, spruce competes with beech only at the very edges. Knowledge of the spatial pattern and the light strategy of both species allows to control the natural regeneration and help in the conversion of spruce monocultures. Indeed, the conversion to a mixed spruce-beech forest requires a combination of various silvicultural practices focused to the light ecology of shade-tolerant beech and semi-shade-tolerant spruce. Beech can regenerate quite easily under a forest canopy (e.g., using a shelterwood system or group selection), whereas for triggering spruce regeneration and its successful growth, a higher light intensity has to be ensured, e.g., by regeneration in the forest edges no deeper than 45 m into the forest or by group felling in areas exceeding 0.04 ha.
Acknowledgements
The research study was supported by the Czech Agency for Agricultural Research (project no. KUS QJ1230330) and by the Mendel University in Brno, Czech Republic (projects no. IGA 84/2013 and no. IGA 33/2014).
References
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Department of Silviculture - Mendel University in Brno - Faculty of Forestry and Wood Technology, Brno, 61300 (Czech Republic)
Corresponding author
Paper Info
Citation
Dobrovolny L (2016). Density and spatial distribution of beech (Fagus sylvatica L.) regeneration in Norway spruce (Picea abies (L.) Karsten) stands in the central part of the Czech Republic. iForest 9: 666-672. - doi: 10.3832/ifor1581-008
Academic Editor
Emanuele Lingua
Paper history
Received: Jan 30, 2015
Accepted: Nov 20, 2015
First online: Mar 12, 2016
Publication Date: Aug 09, 2016
Publication Time: 3.77 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2016
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 53007
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 43104
Abstract Page Views: 3501
PDF Downloads: 4926
Citation/Reference Downloads: 68
XML Downloads: 1408
Web Metrics
Days since publication: 3716
Overall contacts: 53007
Avg. contacts per week: 99.85
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2016): 15
Average cites per year: 1.50
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Growth dynamics of the Norway spruce and silver fir understory in continuous cover forestry
vol. 13, pp. 56-64 (online: 05 February 2020)
Research Articles
Methods for predicting Sitka spruce natural regeneration presence and density in the UK
vol. 12, pp. 279-288 (online: 23 May 2019)
Research Articles
Oak often needs to be promoted in mixed beech-oak stands - the structural processes behind competition and silvicultural management in mixed stands of European beech and sessile oak
vol. 13, pp. 80-88 (online: 01 March 2020)
Research Articles
Short- and long-term natural regeneration after windthrow disturbances in Norway spruce forests in Bulgaria
vol. 11, pp. 675-684 (online: 23 October 2018)
Research Articles
Modelling natural regeneration of Oak in Saxony, Germany: identifying factors influencing the occurrence and density of regeneration
vol. 16, pp. 47-52 (online: 16 February 2023)
Research Articles
Potential relationships of selected abiotic variables, chemical elements and stand characteristics with soil organic carbon in spruce and beech stands
vol. 14, pp. 320-328 (online: 09 July 2021)
Review Papers
Problems and solutions to cork oak (Quercus suber L.) regeneration: a review
vol. 16, pp. 10-22 (online: 09 January 2023)
Research Articles
Understory vegetation dynamics and tree regeneration as affected by deer herbivory in temperate hardwood forests
vol. 10, pp. 837-844 (online: 26 October 2017)
Research Articles
Influence of salvage logging on forest recovery following intermediate severity canopy disturbances in mixed beech dominated forests of Slovenia
vol. 9, pp. 430-436 (online: 07 January 2016)
Research Articles
The impact of seed predation and browsing on natural sessile oak regeneration under different light conditions in an over-aged coppice stand
vol. 9, pp. 569-576 (online: 04 April 2016)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







