
Influence of salvage logging on forest recovery following intermediate severity canopy disturbances in mixed beech dominated forests of Slovenia
iForest - Biogeosciences and Forestry, Volume 9, Issue 3, Pages 430-436 (2016)
doi: https://doi.org/10.3832/ifor1616-008
Published: Jan 07, 2016 - Copyright © 2016 SISEF
Research Articles
Abstract
The practice of salvage logging dead and damaged timber following large high severity disturbances has raised much controversy, partly because of the negative ecological effects that such practices have on forest ecosystems. Many of the studies on salvage logging effects, however, have been done on sites damaged by large, severe disturbances. Less is known about the ecological consequences of salvage logging following intermediate severity disturbances that cause partial canopy damage at smaller scales. We examined the response of the herbaceous layer and tree regeneration to salvaged and non-salvaged treatments following small-scale intermediate severity disturbances in eight mixed beech (Fagus sylvatica L.) dominated forest stands in Slovenia. The cover and diversity of herbaceous vegetation, as well as the density and diversity of tree regeneration were similar between treatments across the study sites. The only notable differences between the treatments were that salvaged sites had a larger proportion of shade intolerant tree species in the regeneration layer, while non-salvaged sites tended to have a more well-developed regeneration layer in taller height classes. The results suggest that salvage logging following small-scale intermediate severity disturbances may not hinder forest recovery in mixed beech dominated forests.
Keywords
Fagus sylvatica, Forest Management, Intermediate Severity, Natural Disturbance, Regeneration, Salvage Logging
Introduction
Salvage logging is often practiced following severe natural disturbances (e.g., fire, windthrow, insect outbreaks), primarily to recover financial losses from damaged timber ([21]). The potential negative effects of such practices on forest ecosystems have raised concerns among ecologists and resource managers. Much of the ecological literature indicates that salvage logging has a negative influence on forest recovery, ecosystem functions, and biodiversity (reviews in [19], [35]). As such, the long-term ecological costs may outweigh short-term economic gains, particularly given that the ecological consequences of salvage logging are poorly understood at longer time scales.
Much of the research that has examined the impact of salvage logging on forest ecosystems has been carried out following very large stand replacing fires, blowdowns, or barkbeetle outbreaks ([3], [20], [12], [13],), often in fire-prone regions or conifer dominated landscapes susceptible to large bark beetle outbreaks. As some authors pointed out, it is not surprising that completely salvaging severely damaged forests over large areas (e.g., 100-1000 ha) would have a substantial impact on forest ecosystems due to the large scale and cumulative impact of severe disturbance followed by salvage logging ([28], [29]).
In many mesic, mixed temperate forests, however, disturbances that create large patches of near stand replacement are rare ([36]), particularly those caused by fire and host-specific insects. In the temperate region of Europe, for example, where Fagus sylvatica is often dominant, forest dynamics is driven by relatively continuous small-scale mortality of canopy trees (i.e., gap dynamics) and periodic intermediate severity damage from wind, ice, or snow associated with more localized storm events ([25], [27]). Such events may remove 20-30% of the canopy at stand scales (e.g. < 10 ha) and create very heterogeneous damage patterns, ranging from small scattered gaps to patches of several thousand m2 ([39], [23]).
Given that intermediate severity disturbances are relatively common, and that salvage logging is widely practiced on both private and public forestland in Europe, understanding the impact of these practices on forest ecosystems is warranted. Very few studies, however, have examined the effects of salvage logging on forest recovery following such disturbances in temperate forests. Peterson & Leach ([28], [29]) compared vegetation recovery in salvaged and non-salvaged sites following intermediate severity windthrow in a mixed forest of Tennessee, USA; while they found clear differences in microsite diversity and abundance between the two treatments, the density and diversity of herbs and tree seedlings did not show a detrimental response to salvaging. Lang et al. ([18]) followed the long-term vegetation response to non-salvage and salvage treatments in a mixed forest damaged by severe wind in Wisconsin, USA. They found that the structure and composition of tree species on both treatments largely converged after 25 years, despite substantial damage to advance regeneration during salvage operations. Working in mixed forests throughout Switzerland, Kramer et al. ([16]) examined the long-term response of tree regeneration to salvage and non-salvage treatments in large windthrow gaps. They found that the density and diversity of tree regeneration did not differ between treatments, presumably because salvage operations did not damage advance regeneration of late successional species, which dominated the forest recovery.
Here we focus on the influence of salvage logging on forest recovery following small-scale intermediate severity disturbances in mixed beech dominated forests. Although beech dominated forests are widespread in Europe, very little attention has been given to the cumulative impact of disturbance and salvage logging in this forest type. We compared the structure and composition of regeneration in neighboring salvaged and non-salvaged areas at eight sites located throughout Slovenia. Many studies have focused on regeneration in beech dominated forests in Europe; these studies indicate that beech, one of the most shade tolerant species in the temperate zone, often forms a persistent bank of seedlings and saplings in the forest understorey, such that even after relatively large disturbances it is able to maintain dominance ([2], [24], [27], [16]). The success of beech during the regeneration stage is thought to limit recruitment opportunities of other more light demanding species, such as Acer pseudoplatanus L., A. platanoides Scop., Fraxinus excelsior L., F. ornus L. and Ulmus glabra Huds. that coexist in these forest communities. We therefore hypothesized that release of regeneration that established prior to disturbances will dominate untreated areas, while salvaged areas will provide opportunities for recruitment of less shade tolerant species due to damage and burial of advance regeneration during logging operations. For the same reason, however, we hypothesized that forest recovery would be more rapid (i.e., taller regeneration) in non-salvaged areas.
Methods
Study area
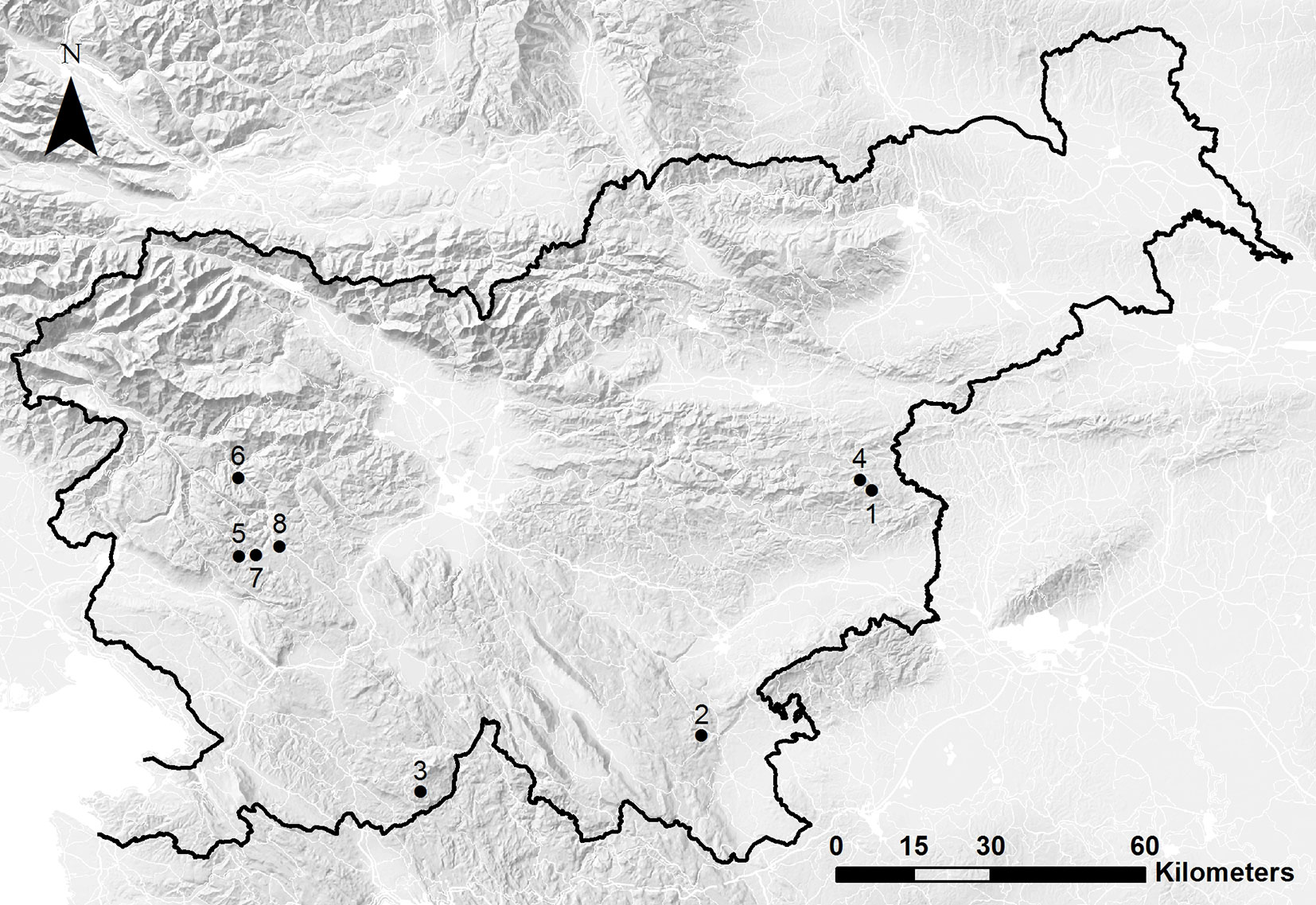
This study was carried out in mixed beech dominated forest stands located throughout Slovenia. Beech dominated forest types make up about 70% of Slovene forest lands and are managed with continuous cover forestry practices that rely on natural regeneration. Salvage logging is almost always practiced following natural disturbances on both public and privately owned forest land. As such, it was difficult to find sites with adjacent salvaged and non-salvaged treatments. After extensive field reconnaissance, we identified 8 sites that included both treatments (Tab. 1, Fig. 1); the main reason for not entirely salvaging these areas were inaccessibility due to rugged terrain and lack of roads and skidding trails. The study sites were mainly damaged by wind disturbances (except two sites damaged by ice and snow) between 1993 and 2008. Sampling was carried out in 2012, such that forest recovery was well underway at the time of sampling. Based on visual estimates from high-resolution aerial photographs provided by the Slovenian Environment Agency, the severity of these disturbances varied between 25-75% canopy removal (Tab. 1).
Tab. 1 - Site characteristics of the eight disturbance areas with adjacent non-salvaged and salvaged treatments. Species are listed according to their abundance in the surrounding stands. (FS): Fagus sylvatica, (OC): Ostrya carpinifolia, (FO): Fraxinus ornus, (FE): Fraxinus excelsior, (AP): Acer pseudoplatanus, (QP); Quercus petraea, (PT): Populus tremula, (CB): Carpinus betulus, (PA): Picea abies, (AA): Abies alba. (SA): Sorbus aria. (lim): limestone, (dol): dolomites, (sil): intrusions of silicates. Disturbance class: (1): 0-25%, (2): 25-50%, (3): 50-75%, (4): 75-100% of disturbed canopy.
| No. | Site | Main tree species |
Bedrock | Year of disturbance |
Slope (°) |
Disturbance type |
Elevation (m) |
Exposure | Size (ha) |
Disturbance Class |
Salvage Technique |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Bohor | FS, OC, FO, SA, AP | dol & lim + sil | 2008 | 34 | wind | 300-460 | S | 5.5 | 2 | tractor |
| 2 | Crmošnjice | FS, AP, QP, PT, CB | dol | 2006 | 32 | ice | 700-830 | NE | 1.3 | 2 | cable crane |
| 3 | Kosmate doline | FS, PA, AP, AA | dol | 1993 | 25 | wind | 1250-1300 | NW-N | 0.7 | 3 | tractor |
| 4 | Lesično | FS, OC, AP, PA, FO | dol & lim + sil | 2008 | 29 | wind | 400-500 | NW | 1.1 | 2 | cable crane |
| 5 | Nemškarica | FS, AA, PA, AP, FE | dol | 2006 | 36 | wind | 610-750 | N-NE | 1.5 | 3 | cable crane |
| 6 | Jagršče | FS, OC, FE, PA, CB | dol | 2007 | 38 | snow | 270-380 | E-NE | 1.5 | 3 | tractor |
| 7 | Zadlog | FS, PA, AP, OC, FE | dol | 2006 | 36 | wind | 700-750 | N | 1.1 | 2 | cable crane |
| 8 | Zala | FS, PA, AA, AP, FE | dol & lim | 2005 | 36 | wind | 400-500 | N-NE | 1 | 2 | cable crane |
Fig. 1 - Map of Slovenia showing the eight study sites. The number of each study site corresponds with those listed in Tab. 1.
Although beech was the dominant canopy species across the study sites, inventory data from the Slovenian Forest Service indicated that a number of other tree species were present in these stands. Those species that were relatively dominant compared to beech included Picea abies (L.) H. Karst. (Norway spruce), Acer pseudoplatanus L. (sycamore maple), Ostrya carpinifolia Scop. (Hop Hornbeam), Abies alba Mill. (European silver-fir), Fraxinus excelsior L. (ash), Quercus petraea (Matt.) Liebl. (Cornish oak), Fraxinus ornus L. (manna ash), Sorbus aria (L.) Crantz (Whitebeam) and Carpinus betulus L. (European hornbeam).
Salvage operations were carried out in the same year or one year following the disturbances with motor-manual cutting and either tractor or cable crane skidding. Both skidding techniques damaged regeneration via removal of tree boles, while tractor skidding included additional damage depending on the surface area of skidding trails at a given site. However, of the three sites harvested with tractor skidding, the Jagršče site was the only site where a new skidding trail was constructed; the other two sites used existing infrastructure.
Field sampling
Within each of the broader disturbance areas, which often included scattered patches of damage within stands, we limited our sampling to disturbed patches that had adjacent non-salvaged and salvaged treatments. These patches ranged from 0.7 to 5.5 ha (Tab. 1). Each treatment within these patches was stratified into 4 approximately equal sized areas and a 100 m2 sample plot was placed at a random location within each area. In each plot, we visually estimated the per cent cover of the herbaceous layer and tree regeneration following the Braun-Blanquet approach, i.e., regeneration cover estimates were divided into herb (< 0.5 m tall) and shrub (0.5-6 m tall) layers - [22]). Additionally, we counted the number of woody stems for each tree species in the following height classes: (i) 20-50 cm; (ii) 51-130 cm; (iii) 131 cm of height up to 5 cm diameter at breast height (dbh); and (iv) dbh > 5cm. This last class included pole size trees typically between 5 and 10 cm dbh, which occurred at low densities in some plots. For each woody stem we categorized browsing damage in 3 classes: (i) up to 10% of lateral shoots browsed; (ii) terminal shoot browsed and/or <50% of lateral shoots browsed; and (iii) terminal shoot and majority of lateral shoots browsed. As an additional indicator of regeneration recovery, we also measured the height of the five tallest individuals in the regeneration layer present in the plot. Finally, we recorded environmental conditions in each plot, including elevation, slope aspect and inclination, relief (i.e., flat, convex, concave), rockiness (% cover), skidding trails (% cover), coarse woody debris (% CWD cover), and erosion (% cover of eroded surface).
The effects of treatment and site factors (elevation, slope, CWD and rockiness) on vegetation cover, regeneration densities, diversity, and height were analyzed with mixed-effects models. These models allow nested error structures as the sample plots were nested within eight sites. Models for tree regeneration density were built with generalized mixed-effects models (GLMMs - [8]) using a negative binomial distribution, while models for vegetation cover and diversity were built with linear mixed-effects models (LMMs - [30]). To meet the assumptions of normality and linearity variables were transformed if necessary. For model diagnostics, both model confidence intervals of parameters and analyzed sets of graphical summaries proposed by Robinson & Hamann ([33]) and Zuur et al. ([40]) were examined. Data were analyzed using R Version 3.0.2. ([31]).
Results
Vegetation cover and density
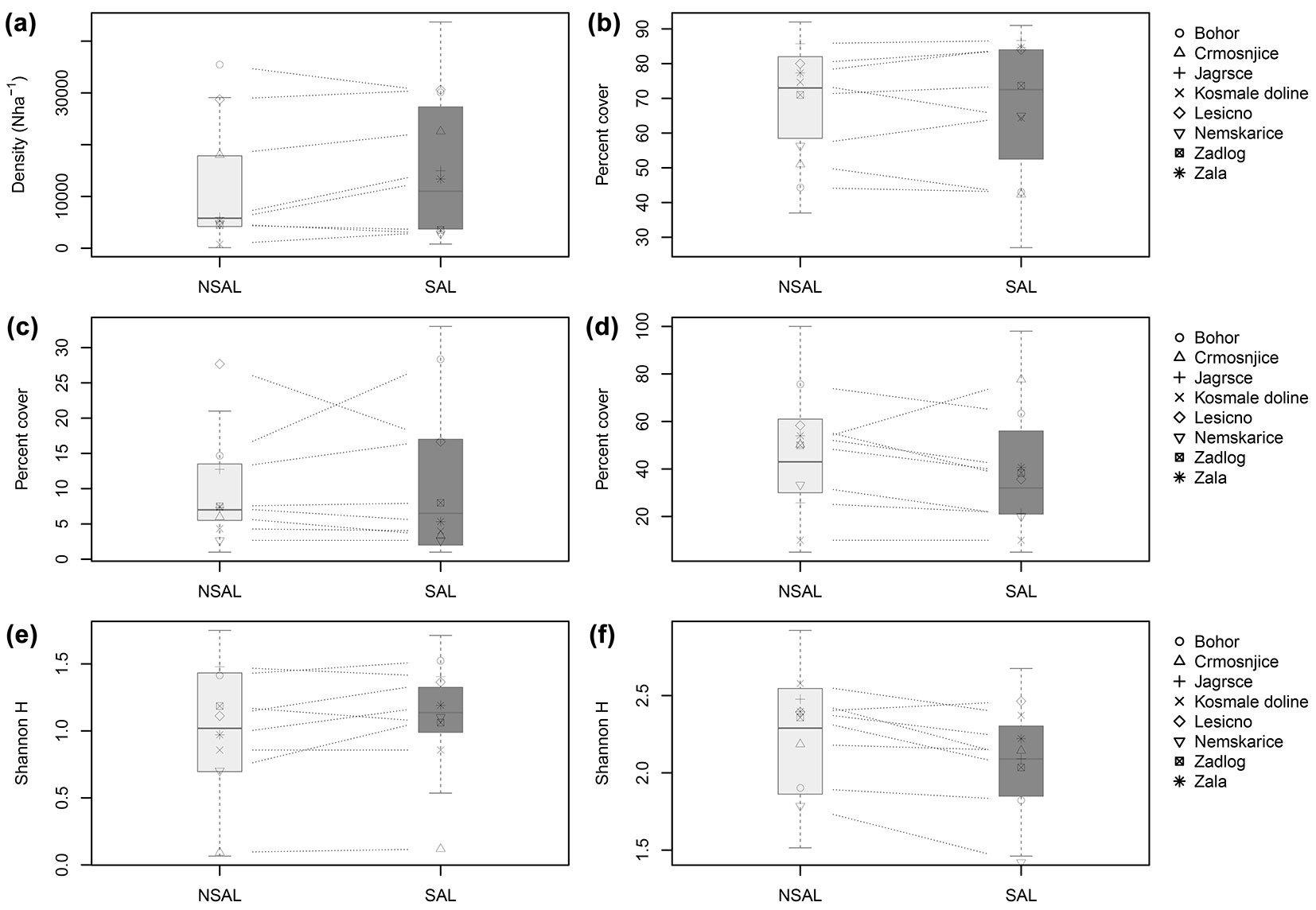
Herbaceous cover and tree regeneration density varied among the 8 study sites. For example, total regeneration density (for both treatments and height classes combined at each site) ranged from 767 to 35467 stems ha-1, respectively. There was, however, a significant decrease in regeneration density with elevation (p = 0.020). With respect to the salvaged and non-salvaged treatments pooled across the 8 sites, the cover of herbaceous vegetation was similar between treatments (p = 0.877 - Fig. 2), while total tree regeneration density (all height classes combined) was higher on salvaged sites, though this difference was not significant (p = 0.059 - Fig. 2). A Spearman’s bivariate analysis revealed that regeneration density was also negatively related with CWD cover (ρ = -0.29), but this was not significant in the GLMM model. The average cover of CWD was 19% in non-salved and 6% in salvaged treatments (p < 0.001). Evidence of erosion was minimal, with an average cover of 4% in both treatments. We found significant differences (p = 0.009) in shrub layer coverage between sites that were harvested with tractor (about 20%) and those with cable crane (about 40%).
Fig. 2 - Box plots of: (a) tree regeneration density (all height classes combined); (b) herb per cent cover; (c) tree and shrub cover in the herb layer; (d) tree and shrub cover; (e) tree regeneration diversity; and (f) herb diversity in non-salvaged and salvaged treatments for the eight study sites combined. The symbols and dashed lines depict individual sites.
Vegetation composition and height structure
A total of 237 herbaceous species and 25 tree species were encountered across the 8 sites, but there were fewer species that dominated each site. For tree species, there were typically about 4-6 species that made up > 90% of the individuals across the sites. The Shannon index for herbaceous diversity was marginally higher on non-salvaged sites (p=0.023), while for trees species (for all height classes combined) it was higher on salvaged sites, though not significant (p=0.200 - Fig. 2).
We documented considerable variability in the height structure of tree regeneration by species across the sites and treatments, patterns which are useful for interpreting successional pathways (Fig. 3). To simplify our interpretation, we divided tree species into three shade tolerance categories based on Ellenberg’s indicator values (EIV) for light ([6]): (i) tolerant (EIV < 4); (ii) intermediate (EIV = 4); and intolerant (EIV > 4). Contrary to our expectations, shade tolerant species, namely Fagus sylvatica, Carpinus betulus, and Abies alba, were not dominant across the study sites (44% of stems), nor were they clearly dominant in non-salvaged sites (47% in non-salvaged versus 41% in salvaged - Fig. 3, Fig. 4). However, they were more abundant in taller regeneration height classes (60% of the two tallest height classes). There were no significant differences between treatments with regard to either the combined cover (p=0.072) or density (p=0.110) of these three shade tolerant species. It is worth noting, however, that these shade tolerant species made up a higher proportion of the regeneration layer in non-salvaged treatments, although this was not significant (Fig. 4).
Fig. 3 - Density of tree species in the regeneration layer by height class and treatment. Only the six most abundant tree species, which made up > 90% of the total stems, are shown for each site.
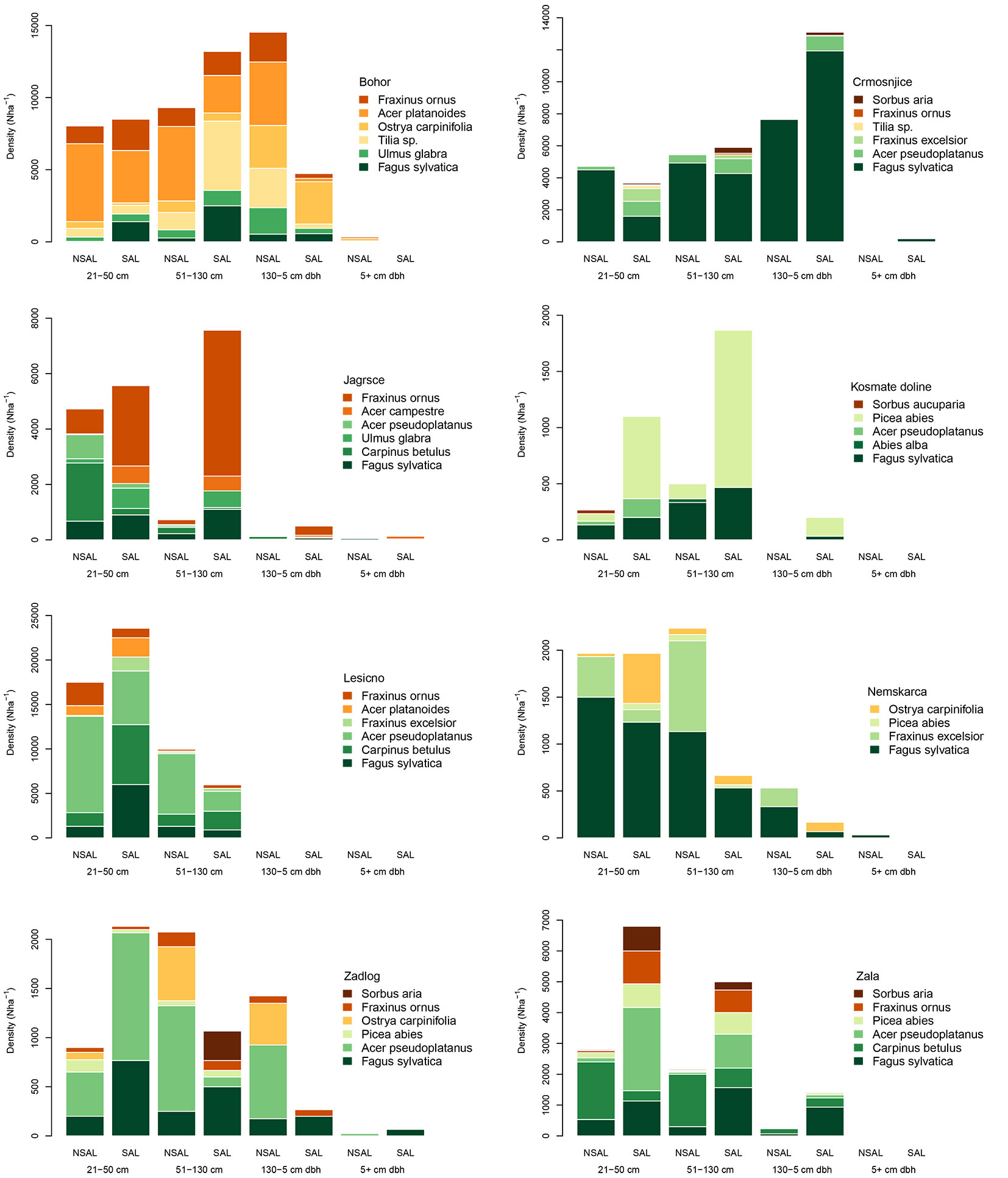
Fig. 4 - Box plots showing the proportion (%) of (a) shade tolerant, (b) intermediate, and (c) shade intolerant tree species in the regeneration layer by treatment. The symbols and dashed lines depict individual sites.
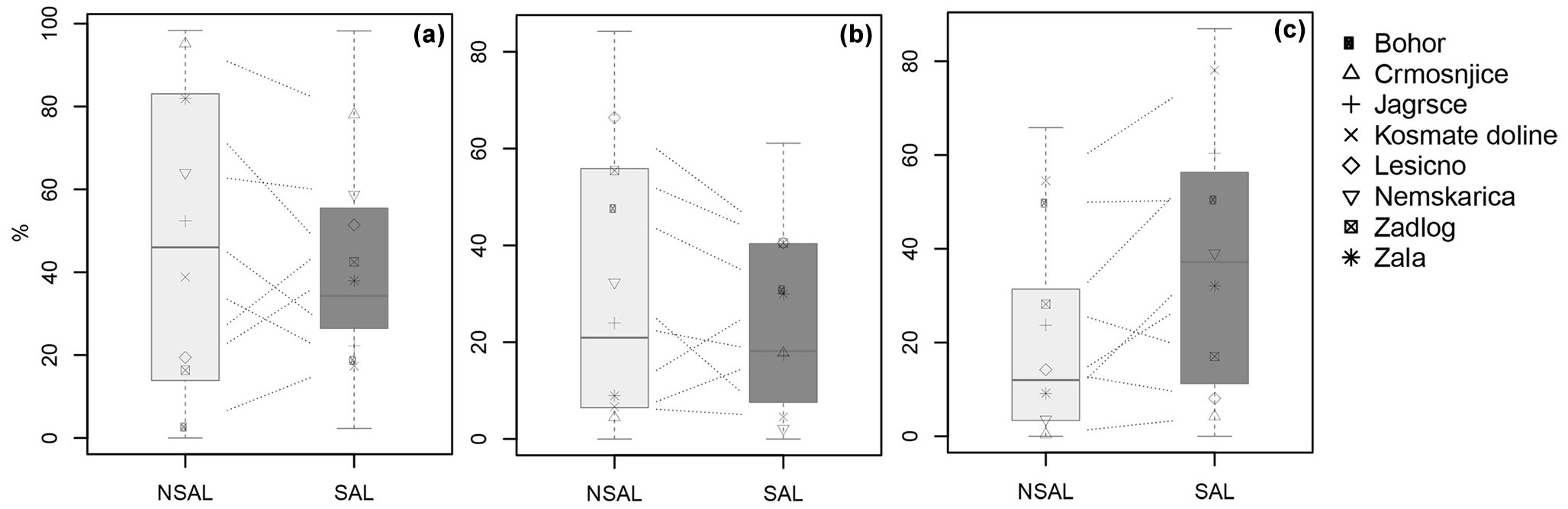
Less shade tolerant tree species, especially species like Acer pseudoplatanus, A. platanoides, Fraxinus excelsior and F. ornus, were equally or more abundant than shade tolerant species across the height classes, but the patterns between the salvaged and non-salvaged treatments were variable (Fig. 3). At some sites, there was a general trend toward higher densities of less shade tolerant species in the salvaged treatment (e.g., Crmošnjice, Zala), while the opposite pattern was found on other sites (e.g., Zadlog). Nevertheless, we did find that shade intolerant species made up a larger proportion (p= 0.013) of the regeneration layer in salvaged (36 %) compared to non-salvaged sites (23 % - Fig. 4). This was consistent across all height classes. The proportion of tree species with intermediate shade tolerance was similar between treatments, but decreased with increasing height classes in both treatments.
There was also no clear pattern with regard to the overall height structure of the regeneration layer between treatments. On sites with a well developed layer of regeneration in the H > 131 cm and dbh < 5 cm class, for example, some sites had a higher overall density in non-salvaged treatments (i.e., Bohor, Nemškarica, Zadlog), while others had a higher density in the salvaged treatment (i.e., Crmošnjice, Zala). Overall, stems in the two smaller classes combined had a higher density in salvaged treatments (p = 0.038). Likewise, the Braun-Blanquet cover estimates indicated that the combined cover of the moss and herb layer was higher in salvaged sites (78%) when compared to non-salvaged (72%), while the combined cover of the shrub and tree layer was higher in non-salvaged sites (46% vs. 40%); none of these differences were statistically significant (p = 0.170, p = 0.164). The overall mean height of the tallest stems was marginally taller in non-salvaged (189.0 cm; n=106) compared to salvaged treatments (167.1 cm; n=95 - p=0.060). Browsing damage on regeneration was relatively low across the sites (80 % of the regeneration was classified in the category with < 10% of lateral shoots browsed) and significantly higher (χ2, p=0.000) on salvaged sites.
Discussion
While we found substantial variability in vegetation structure and composition among sites and between salvaged and non-salvaged treatments within individual sites, we did not identify compelling differences between salvaged and non-salvaged treatments pooled across the study sites. Salvaged treatments on many of the study sites had marginally higher species diversity in the regeneration layer and a higher proportion of shade intolerant tree species than in non-salvaged treatments, yet there was no clear evidence that release of shade-tolerant advance regeneration dominated non-salvaged treatments. Therefore, we found limited support for our first hypothesis. Our second hypothesis, that salvaged treatments would be less advanced in their development due to damaged advance regeneration, also received only partial support. On the whole non-salvaged treatments had marginally higher cover, density, and heights within the taller regeneration classes.
Although we do not have data on the pre-disturbance forest structure and composition, national forest service inventory plots indicate that beech was dominant in the canopy across most of the sites (60-90% of canopy layer), except Kosmate doline, where spruce was co-dominant. The higher tree species diversity found in the study areas (relative to their abundance in the canopy), regardless of treatment type, suggests that increased light levels following the natural disturbance events facilitated recruitment of less shade-tolerant species. Although we cannot distinguish stems that were present as advance regeneration prior to disturbance from those that established afterwards, the height structure data indirectly suggest that both pathways were important for the recovery process. It is also important to note that the study areas varied in time since disturbance, yet disturbances in all but one site (Kosmate doline - a cold, high elevation site with slower development) were within a four year span, such that recovering forests were likely within similar stages of development.
Several reasons may explain the lack of a clear treatment effect. First, given that we focused on intermediate severity wind disturbances that damaged relatively small areas, it is reasonable to assume that much of the soil, herbs, and advance tree regeneration were left intact, except for vegetation crushed by fallen trees. Furthermore, the salvage operations were also relatively moderate and relied on existing skidding trails and forest roads, such that much of the soil and vegetation on these sites likely remained relatively undamaged. This is well exemplified by the low cover of eroded bare soil across the sites, which averaged only 4% in both treatments. In a large study of damage to regeneration resulting from tractor skidding and cable crane harvesting across 51 stands in Slovenia, the average amount of damaged regeneration was 21 and 16%, respectively ([15]). This suggests that both extraction techniques in this study (tractor skidding and cable crane) were unlikely to have caused a substantial amount of residual stand damage. In contrast, harvester machines, which are often employed for large-scale salvage operations, drive over a larger portion of a given stand, which could result in greater residual damage to soils and regeneration ([37]).
A second reason may be that environmental differences between neighboring treatments were more important drivers of vegetation recovery than treatment. Because it was difficult to find study sites with neighboring salvaged and non-salvaged treatments throughout beech forests in Slovenia, we were unable to control for variation in site conditions between treatments, such as small differences in slope aspect and steepness or soil conditions, which were likely to confound treatment effects. For example, in a large-scale study on the influence of salvage logging on forest recovery across Swiss forests, Kramer et al. ([16]) found that site factors (i.e., soil pH and ground vegetation) were more important than treatment.
Although we did not find substantial differences in vegetation structure and composition between treatments, the trends in the pooled data set provide some support for our predictions. Both the density and diversity of tree regeneration were slightly higher in salvaged treatments, while there was evidence of a more well-developed regeneration layer in non-salvaged areas. It seems likely that removal of coarse woody debris, which may act as a physical barrier to regeneration, coupled with less competition from taller advance regeneration, enabled establishment and recruitment of a wide range of tree species in the salvaged areas. The higher cover of taller stems in the non-salvaged area may be ecologically important because these individuals are likely to form the next canopy layer and may shade out many of the stems in the lower layers.
Differences in the cover and composition of herbs were less clearly related to treatment, perhaps because they respond more to underlying differences in site conditions. A number of studies have indicated that a dense herb layer may develop following disturbance to the forest canopy, which may inhibit tree regeneration during early stages of forest recovery ([38], [12], [14]). The relatively low density of tree regeneration across the study sites, with an overall average of 13770 stems ha-1, provides indirect evidence of this. For comparison, published regeneration densities from unmanaged beech dominated forests in Europe, where smaller scale gap dynamics drive regeneration dynamics, are often substantially higher ([4], [34], [11]).
One of the more surprising findings was the low level of browsing damage to regeneration across the study sites. The density of red deer in Slovenia is high, with more than 10 deer km-2 in some regions ([26]). Moreover, compared to species of Fagus sylvatica and Carpinus betulus regeneration, many of the less shade tolerant tree species in the regeneration layer, particularly Acer pseudoplatanus, A. platanoides, Ulmus glabra, Fraxinus excelsior and F. ornus are highly preferred browse species ([10]). We did observe lower levels of browsing in non-salvaged treatments, which some authors attribute to physical barriers provided by downed trees ([32], [1]).
It is difficult to make broad generalizations regarding the influence of salvage logging on forest recovery across forest types and disturbance regimes. While there is clear evidence of the negative effects of salvaging on regeneration in some circumstances (e.g., large-scale high severity disturbance and salvaging - [3], [13]), the evidence is less clear following a combination of intermediate severity disturbance and salvage logging ([28], [18], [5], [16]). It is important to note that we focused on the influence of salvage logging on vegetation in this study. There is overwhelming evidence that salvage logging has a negative influence on a number of other structures and functions of forest ecosystems. For example, salvaging large amounts of dead coarse wood removes key food and habitat for species dependent on these biological legacies ([9], [17]). Given the low volume of dead wood across managed forests in Europe ([7]), refraining from salvage logging some post-disturbance sites should be advised in forests managed for multiple functions, such as many publicly owned forest lands in Europe that integrate both ecological and economic functions. On private land, however, where owners often have a vested interest in recovering value from downed wood following disturbances, our study suggests that salvage logging may not hinder forest development following small-scale intermediate severity disturbances in mixed beech dominated forests.
Acknowledgements
The study was supported by the applied project “Ecological restoration of natural disturbances in forests” (L4-4091) as well as by the Pahernik foundation. We thank students from the Department of Forestry, University of Ljubljana (Slovenia) that conducted the field work. We also thank the Slovenian Forest Service for help with identifying suitable research sites. Finally, we are grateful to Aleš Poljanec who provided the data from the Slovenian Forest Service inventory for the research sites.
References
Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Andrej Rozman
Thomas A Nagel
Jurij Diaci
Biotechnical Faculty, University of Ljubljana, Ljubljana (Slovenia)
Slovenian Academy of Science and Arts, Jovan Hadži Institute of Biology, Ljubljana (Slovenia)
Corresponding author
Paper Info
Citation
Fidej G, Rozman A, Nagel TA, Dakskobler I, Diaci J (2016). Influence of salvage logging on forest recovery following intermediate severity canopy disturbances in mixed beech dominated forests of Slovenia. iForest 9: 430-436. - doi: 10.3832/ifor1616-008
Academic Editor
Emanuele Lingua
Paper history
Received: Feb 24, 2015
Accepted: Aug 27, 2015
First online: Jan 07, 2016
Publication Date: Jun 01, 2016
Publication Time: 4.43 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2016
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 51532
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 42455
Abstract Page Views: 3532
PDF Downloads: 4010
Citation/Reference Downloads: 24
XML Downloads: 1511
Web Metrics
Days since publication: 3783
Overall contacts: 51532
Avg. contacts per week: 95.35
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2016): 15
Average cites per year: 1.50
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Review Papers
Towards better practices of salvage logging for reducing the ecosystem impacts in Mediterranean burned forests
vol. 13, pp. 360-368 (online: 25 August 2020)
Research Articles
The concept of green infrastructure and urban landscape planning: a challenge for urban forestry planning in Belgrade, Serbia
vol. 11, pp. 491-498 (online: 18 July 2018)
Research Articles
Communicating spatial planning decisions at the landscape and farm level with landscape visualization
vol. 7, pp. 434-442 (online: 19 May 2014)
Research Articles
Post-fire effects and short-term regeneration dynamics following high-severity crown fires in a Mediterranean forest
vol. 5, pp. 93-100 (online: 30 May 2012)
Technical Advances
Forest planning and public participation: a possible methodological approach
vol. 5, pp. 72-82 (online: 12 April 2012)
Research Articles
Can traditional selective logging secure tree regeneration in cloud forest?
vol. 10, pp. 369-375 (online: 07 March 2017)
Review Papers
Public participation: a need of forest planning
vol. 7, pp. 216-226 (online: 27 February 2014)
Research Articles
Public participation GIS to support a bottom-up approach in forest landscape planning
vol. 6, pp. 347-352 (online: 29 August 2013)
Research Articles
Role of serotiny on Pinus pinaster Aiton germination and its relation to mother plant age and fire severity
vol. 12, pp. 491-497 (online: 02 November 2019)
Review Papers
Remote sensing of selective logging in tropical forests: current state and future directions
vol. 13, pp. 286-300 (online: 10 July 2020)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







