
Differences in early dynamics and effects of slope aspect between naturally regenerated and planted Pinus sylvestris woodland on inland dunes in Poland
iForest - Biogeosciences and Forestry, Volume 9, Issue 6, Pages 875-882 (2016)
doi: https://doi.org/10.3832/ifor1728-009
Published: Jun 17, 2016 - Copyright © 2016 SISEF
Research Articles
Abstract
There is little knowledge of the effects of landform relief on early growth dynamics and competitive interactions of Pinus sylvestris (Scots pine) stands on inland dunes, which could potentially be substantial. The goal of this study was to examine and compare early dynamics (based on growth parameters and properties of the understorey vegetation) and the effects of slope aspect in naturally regenerated and pine-planted woodland on inland dunes in northern Poland. Growth parameters, tree density, and understorey vegetation were monitored on north- and south-facing slopes in a 26.3 ha post-fire area with natural secondary succession and eight even-aged pine-planted stands, 5-34 years old. Clear differences were detected between the woodland types, in both growth parameters of pines with similar ages and effects of slope aspect on the pines. In the natural regeneration area north-facing slopes provided favorable conditions for natural encroachment by pines. Tree density was higher, and the pines were taller and thicker, on north-facing than on south-facing slopes. In contrast, in the pine-planted area pines were larger on south-facing slopes, although growth conditions were less favorable than on north-facing slopes. However, in the pine-planted area the north-facing slopes had significantly higher herb cover, dominated by Deschampsia flexuosa, indicating that even the presence of a relatively low grass species can impede early growth of Scots pine. The understorey species composition differed in the natural regeneration area, being dominated by Calluna vulgaris on north-facing slopes and Corynephorus canescens on south-facing slopes. The results reveal that interactions between landform, natural dynamics, planting practices, and competitive interactions in woodlands on inland dunes are complex, and should be considered in efforts to manage them efficiently.
Keywords
Scots Pine, Growth Dynamics, Dunes, Slope Aspect, Forest Succession, Pine Plantations, Sandy Areas, Podzols
Introduction
Scots pine (Pinus sylvestris) is the most widely distributed species of the Pinaceae family, and a commercially important forest tree in large parts of its native range. Due to its high stress tolerance, it is also widely planted to stabilize and restore habitats in sites with poor conditions, for example, to fix dunes with dry, poor sandy soil ([30], [9]). However, young pine stands are often severely threatened by extreme stress or disturbance in such environments ([49], [50], [25]). Thus, thorough understanding of the biotic and abiotic processes in Scots pine forests is needed, particularly those in “high risk environments” during early development stages, when the young trees are most susceptible to stresses and disturbances. In early development phases, young trees and understorey vegetation (shrubs and herbaceous weeds) strongly compete for the nutrients, water and light required for their growth and survival, in both natural woodlands during succession ([31], [11], [29]) and managed forests ([2], [35]). The competition is most evident under extreme conditions, e.g., when soil moisture is generally low ([45], [24]), a typical characteristic of the dry, sandy soils of inland dunes ([50], [20], [34]). Fire can help tree species compete with understorey vegetation ([3]), and reportedly facilitate natural regeneration of Scots pine ([19], [23]). However, fires can also impair Scots pine’s encroachment of a new ecosystem. For example, slow-moving fires can increase the temperature of Scots pine seeds to over 90 °C and harm their germinative capacity, thus helping competitor species ([26]).
Landform relief is a major factor in plant growth and dynamics, as it determines both the gradient and aspect of slopes, and thus the amount of solar radiation which strongly influence moisture and temperature regimes ([8], [47], [34]). However, as slope aspect has the strongest effects on vegetation in large-scale relief forms, previous research on relief’s effects on plant growth has mainly focused on mountainous areas ([27], [16], [32], [7], [40], [6]). There has been less investigation of its effects in dune environments, which have relatively slight relief, and most of this investigation has focused on coastal dune ecosystems ([30], [1], [42], [25], [9]). Some slope-related variations in the vegetation and microclimate of inland dune environments have been reported ([20], [34]), but more detailed observation and analysis of the mechanisms responsible for the observed variations are required.
In the northern hemisphere, north-facing slopes receive less solar radiation than south-facing slopes, and thus generally have a more humid microclimate, more fertile soil, and lower temperatures ([8], [13], [34], [35], [39]). Hence, aspect also strongly influences the species composition of trees, shrub communities, and herb layers ([47], [20], [34]). Furthermore, it may significantly affect timber production; notably, many studies have shown that stands grow faster on north-facing slopes than on south-facing slopes in northern temperate forests due to their higher soil moisture and fertility ([27], [16], [40]).
Ecological approaches are increasingly applied in land restoration programs and forest management, and it has been suggested that silvicultural practices should reflect the dynamics of naturally developing ecosystems ([38]). A potentially valuable way to identify practices that may be suitable for such strategies is to compare processes in natural woodlands and managed forests ([31]). As most inland dune habitats of Central Europe are currently covered by even-aged, planted Scots pine monocultures, extensive areas with natural vegetation are scarce, and the only such area remaining in Poland is the Torun artillery training ground, in the northern part of the country.
The objectives of this study were to examine and compare early dynamics (based on growth parameters and properties of the understorey vegetation) and effects of slope aspect in naturally regenerated and pine-planted woodland on inland dunes in Poland. It was hypothesized that early dynamics would differ between the two woodland types, that slope aspect would affect the dynamics, and that the results would provide valuable insights for managing forests and reforesting dune environments.
Materials and methods
Site description and experimental design
The study was conducted at the Torun artillery training ground (52° 55′ N, 18° 36′ E - Fig. 1), located in one of the largest inland dune fields of Poland. The dominant soils here are sandy, acidic, dry and nutrient-poor Podzols ([4], [34]). The mineral material of the soil has very homogeneous composition and texture, being highly dominated (85-99%) by quartz ([20]), and approximately 80% of the particles are in the 0.1-0.25 mm fraction ([34]). The study area has a transitional type of climate, between oceanic and continental. The mean annual temperature is 7.9 °C, with monthly means ranging from -2.2 to 18.1 °C in January and July, respectively. The average annual precipitation is 522.5 mm, July being the wettest month. The mean length of the annual growing season at the study area is 218 days ([46]).
Fig. 1 - Location of the study area.
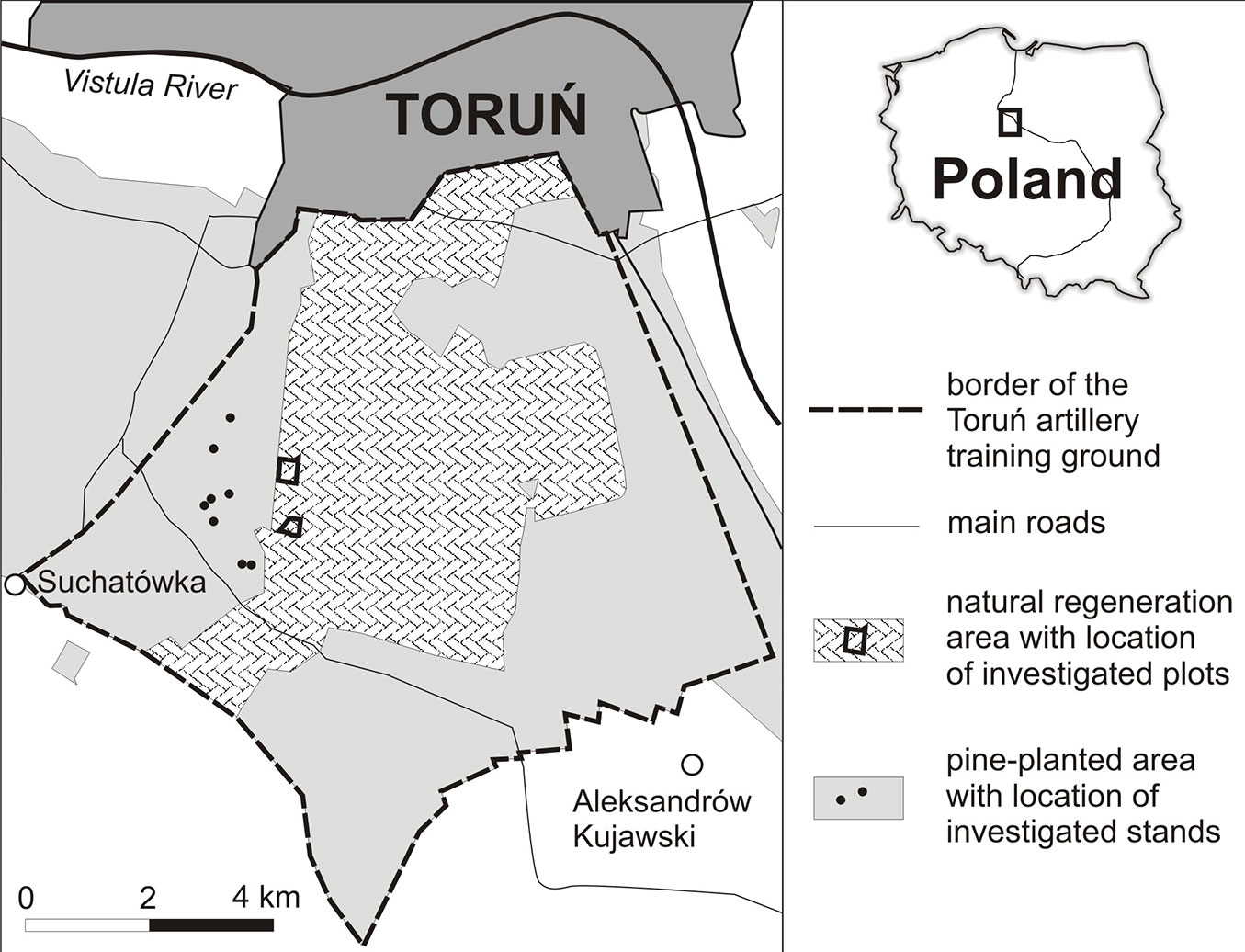
For the comparative study two plots were established in a natural regeneration area (NRA) and pairs of plots in eight stands in a pine-planted area (PPA). The two NRA plots, covering 26.3 ha in total and spaced approximately 1 km apart (Fig. 1), were established for inclusion in a long-term research program in 2011, when all trees in the plots at least 1 m tall were measured ([37]).
The studied NRA had been deforested for military purposes in the 1950s, and since then the main factors influencing development of the vegetation had been natural secondary succession and fires that had periodically retarded the succession. As the plots were in a military area, no forest management practices (soil preparation, thinnings etc.) had been applied to either of them for decades. The last two fires in the NRA (both of which burned the two study plots) occurred in 1989 and 1991. The fires were not very severe as there had been relatively little fuel to burn, because in the late 1980s only about 30 years had passed since deforestation in the 1950s. Thus, the vegetation in the studied plots was similar to the vegetation observed in 2011. Nevertheless, the fires eliminated most of the trees on both plots, and in 2011 only eight pre-fire pines were present in the whole 26.3 ha study area. The oldest two of these trees were 41 and 45 years old, and the other six were younger than 30 years. When the measurements were taken in 2011 the plots were covered by stands with a loose tree canopy dominated by pines (Fig. 2) with ages quite evenly distributed across the range 0-22 years (Tab. 1), reflecting the gradual encroachment of trees in the NRA in the years following the last fires. As the potential natural vegetation on the studied dunes is subcontinental Peucedano-Pinetum forest ([10]), Pinus sylvestris accounted for most (82%) of the trees encroaching in the plots. Silver birch (Betula pendula) accounted for another 17%, and percentages of other species were minimal ([37]).
Fig. 2 - A study plot located in the natural regeneration area (NRA). Stands that are part of planted and managed forests can be seen in the background.

Tab. 1 - Age distribution of pines in the natural regeneration area (NRA).
| Age classes (years) |
No. of pines |
|---|---|
| <10 | 316 |
| 11-15 | 189 |
| 16-20 | 176 |
| 21-25 | 13 |
| 26-30 | 4 |
| >30 | 2 |
The PPA plots were established in the external zone of the Torun artillery training ground, where the forest was divided into even-aged sections of P. sylvestris stands representing different age classes. The stands were usually pine monocultures with small proportions of other species, mainly B. pendula. Unlike forest in the NRA, stands in the PPA had been subjected to standard forest management practices. Eight PPA stands were selected to cover the age range of most pines growing in the NRA (5, 6, 7, 10, 14, 17, 22 and 34 years old - younger stands were excluded, because pines in them were much shorter than 1 m). All of the PPA stands selected for inclusion in the study had replaced at least one previous generation of clear-cut forest, had not been exposed to fire for at least 50 years, and were located on a dune with a south-facing and a north-facing slope, both of which had received identical silvicultural treatments since planting.
All of the study plots, representing both woodland types, were located within 3 km of each other (Fig. 1). They were situated beside army training grounds, and vegetation development had not been impacted by military activities (e.g., explosions etc.) in recent decades. The mean relative height of the investigated dunes is about 8 m. The only source of water for the plants on the dune landforms is precipitation, as no groundwater was available to the roots.
Data collection and analysis
In both NRA plots the GPS location, total height (H) and diameter at 5 cm above ground level (D) of all pines at least 1 m tall (700 in total) were measured in autumn 2011. Tree height was determined using a calibrated pole or a Vertex IV hypsometer (Haglöf, Långsele, Sweden), as appropriate, and the diameter using a caliper. In addition, the height of the lowest living branch (H1) of 177 pines representing the full-size range of trees included in the study was measured, and their ages were determined by counting the tree-rings in cores extracted with an increment borer. To determine whether trees were on a north-facing or a south-facing slope the GPS positions of NRA pines were transferred to ArcGIS® 9.3 (ESRI, Redlands, CA, USA). North- and south-facing slopes were defined as polygonal areas with azimuths of 315-45° and 135-225°, respectively, and a minimum inclination of 5°. In total, 112 such polygons were identified, 60 representing north-facing slopes and 52 representing south-facing slopes, supporting 180 and 69 pines, respectively (Tab. 2).
Tab. 2 - Characteristics of natural regeneration area polygons, obtained with ArcGIS, representing slopes with northerly and southerly aspects. (SD): standard deviation.
| Variable | Slope aspect | |
|---|---|---|
| Northerly | Southerly | |
| Number of polygons | 60 | 52 |
| Mean (± SD) area of a polygon (m2) | 526 ± 639 | 1122 ± 3240 |
| Mean (± SD) number of pines in a polygon | 3.0 ± 5.2 | 1.3 ± 3.5 |
| Total number of pines in all polygons | 180 | 69 |
To obtain tree measurements in the PPA stands 16 rectangular plots were established, two in each stand, one on a north-facing slope and the other on a south-facing slope (Tab. 3). As silver birch strongly affects the growth of surrounding trees ([48]), plots were only established in parts of the stands where the only trees were Scots pines; consequently, the plot area varied from 48 to 92 m2, due to restrictions imposed by the presence of silver birch. In 2013, the H, H1, and D of all the pines in the plots (623 in total) were measured.
Tab. 3 - Characteristics of stands in plots established in the pine-planted area (PPA).
| Age of stand (years) |
Slope aspect |
Plot area (m2) |
Number of pines |
Tree density (pines ha-2) |
|---|---|---|---|---|
| 5 | Northerly | 66 | 44 | 6687 |
| Southerly | 58 | 39 | 6667 | |
| 6 | Northerly | 50 | 34 | 6800 |
| Southerly | 50 | 32 | 6400 | |
| 7 | Northerly | 90 | 55 | 6119 |
| Southerly | 90 | 60 | 6675 | |
| 10 | Northerly | 81 | 55 | 6834 |
| Southerly | 92 | 52 | 5662 | |
| 14 | Northerly | 50 | 33 | 6571 |
| Southerly | 49 | 32 | 6492 | |
| 17 | Northerly | 57 | 31 | 5366 |
| Southerly | 48 | 25 | 5164 | |
| 22 | Northerly | 74 | 33 | 4478 |
| Southerly | 82 | 31 | 3758 | |
| 34 | Northerly | 174 | 32 | 1839 |
| Southerly | 190 | 35 | 1842 |
Using the initial measurements, the slenderness (H/D ratio) and relative crown length (RCL, obtained by dividing the crown length, H-H1, by H then multiplying by 100), were calculated for all the measured pines in both woodland types. Moreover, to avoid drilling increment bores in all pines in the NRA, the relationships between age and both height and diameter were calculated by linear regression. Age was found to be more strongly related to diameter (R2=0.84) than height (R2=0.81), so the ages of pines that were not drilled in the NRA plots were estimated (in years) using the equation (eqn. 1):
where D is the diameter (in cm) taken at 5 cm above the ground level.
The understorey vegetation of both woodland types was also investigated. In the PPA stands a plot was established in the middle part of each studied slope, while in the NRA six plots were established on both north-facing and south-facing slopes. All plots were 200 m2 in area and there were 28 in total, 12 in the NRA and 16 in the PPA. In each plot herb cover was measured as a percentage and vascular species composition was determined by the Braun-Blanquet method ([5]). All of this work was performed during the 2013 growing season.
Environmental conditions were indirectly assessed by a bio-indication technique based on the weighted abundances of specific plant species as indicators of positions along environmental gradients ([15]). Such approach has been extensively used in ecological studies ([22], [18]). Using indicator values proposed for the vascular plants of Central Europe by Ellenberg et al. ([15]), indices were calculated for the ecological variables light (L), temperature (T), continentality (K), moisture (F), soil reaction (R) and nitrogen (N) for each of the 28 plots where understorey vegetation properties were investigated. Salinity was excluded as it is only relevant for saline soils, which were not present at the study site. The latin names of plant species follow Flora Europaea ([43]).
Statistical analysis
Non-parametric tests were used to test for differences in mean values of parameters between groups. Differences in measured variables of pines growing on north- and south-facing slopes were evaluated using the Mann-Whitney U test. Pooled data for all of the pines in the 60 polygons on north-facing slopes and 52 polygons on south-facing slopes in the NRA were used for this comparison, but in the corresponding PPA comparison pines on north- and south-facing slopes in each stand were compared separately, as these even-aged stands differed in age. The Mann-Whitney U test was also used to assess the difference in tree density between north- and south-facing slopes in the NRA, but not the PPA, where the tree density was strongly influenced by the forest management practices, in addition to natural factors. Furthermore, it was used to assess the difference in mean herb cover between plots on six north-facing and six south-facing slopes in the NRA, and between plots on north-facing and south-facing slopes in the eight focal stands in the PPA. The Kruskal-Wallis test was used to assess the significance of differences in the Ellenberg indicator values for all 28 plots where the understorey vegetation was characterized. The statistical analyses were performed using the software package STATISTICA® version 9.0 (StatSoft, Tulsa, OK, USA). Detected differences were deemed significant if P < 0.05, and errors indicated in the text with ± symbols after mean values are standard deviations.
The differences in understorey vegetation species composition among the 28 plots were also analyzed by ordination techinques using the CANOCO package ver. 4.5 ([41]), after transforming cover values obtained for species with the Braun-Blanquet scale into ordinal scale values as follows: r→1, +→2, 1→3, 2→5, 3→7, 4→8, 5→9 ([44]). Next, the length of the environmental gradient for plant occurrence was determined by Detrended Correspondence Analysis (DCA). The length was 3.846 SD (standard deviation) units and the pattern of vascular plant composition was evaluated by Principal Component Analysis (PCA - [41]).
Results
Comparison of pine growth parameters in the two woodland types
There were clear differences in pine dimensions between the woodland types, which became more pronounced with increases in age. The parameters of young pines up to 7 years old were similar at both sites, but as the trees aged, they diverged (Fig. 3). Furthermore, pines in PPA stands tended to be taller but thinner (thus having a higher slenderness ratio, H/D) than those of corresponding ages in the NRA (as also shown in Fig. 3).
Fig. 3 - Growth parameters of pines in the two studied woodland types. (H): total height; (D): diameter at 5 cm above ground level; (H/D): slenderness; (RCL): relative crown length. The solid lines and circles indicate data for the natural regeneration area while dotted lines and squares indicate data for stands in the pine-planted area. The vertical lines indicate standard deviation values for the growth parameters of pines in the pine-planted area. R2 values were obtained from linear regression of indicated growth parameters of pines in the natural regeneration area against their age.
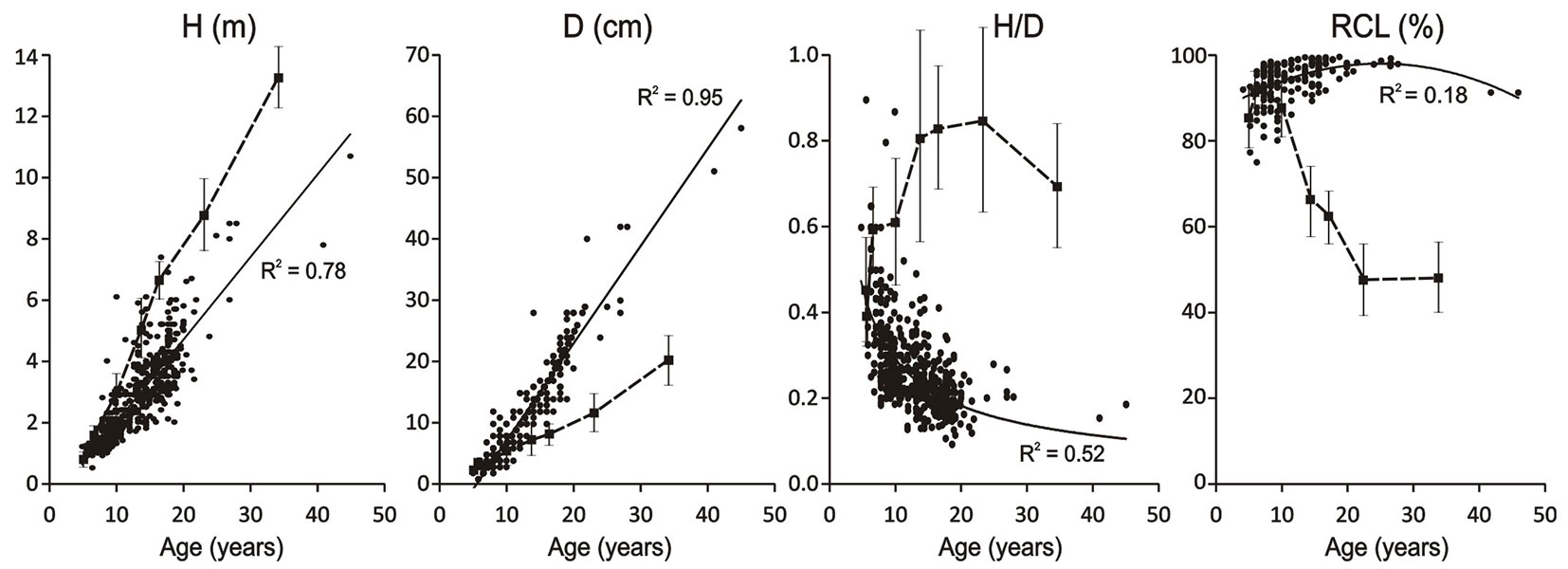
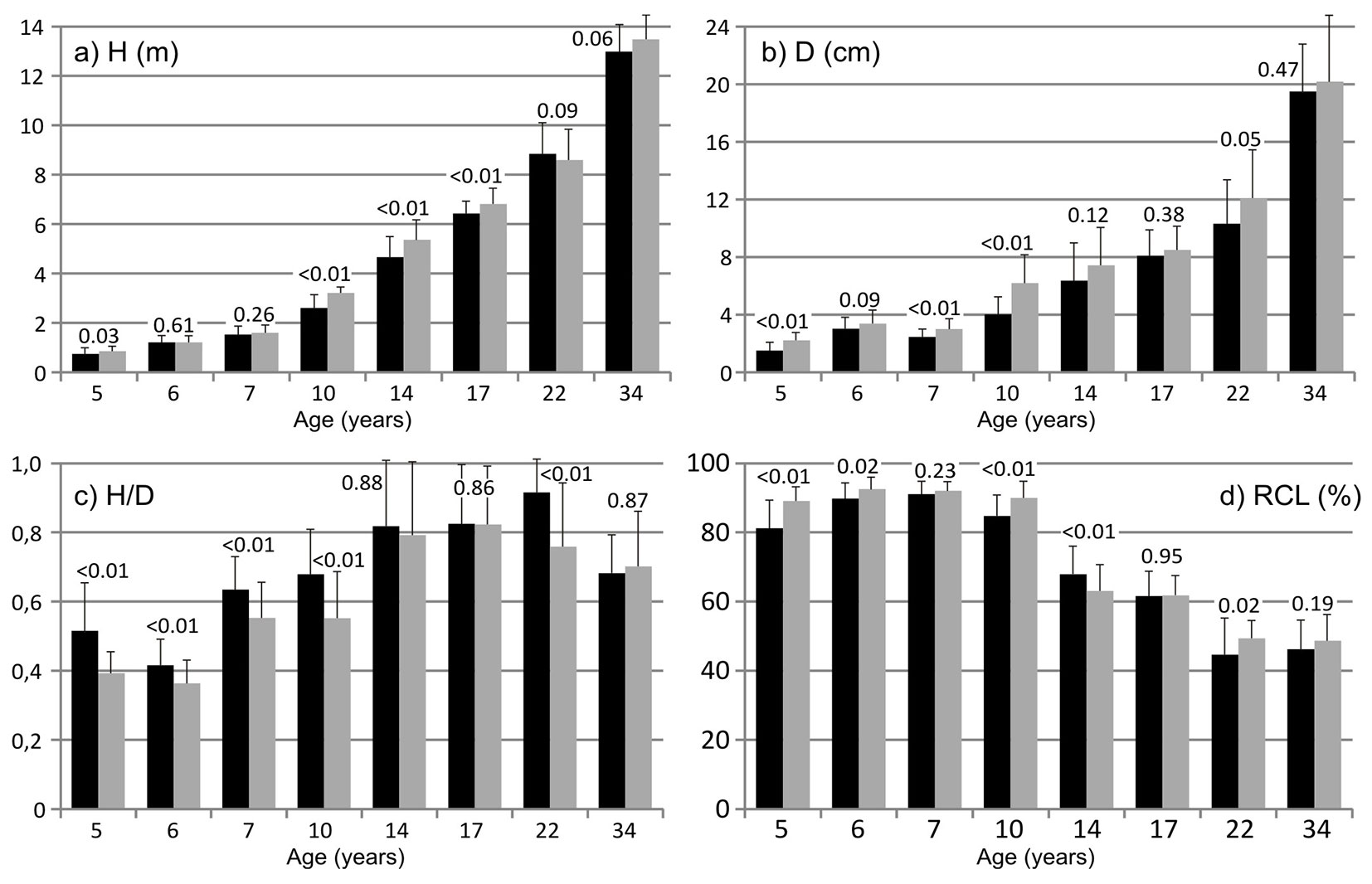
The slope aspect affected the growth dynamics of young pines, in both woodland types, but the effects qualitatively differed and were stronger in the NRA than in the PPA. In the PPA stands, trees growing on south-facing slopes generally had significantly higher mean H and D values than those growing on north-facing slopes (Fig. 4a, Fig. 4b). Furthermore, in five of the eight studied stands, trees on north-facing slopes had higher (P < 0.01) slenderness ratios than trees of the same age on south-facing slopes, while in the other three stands there was no significant between-slope difference in slenderness ratio (Fig. 4c). The only considered growth parameter that was not significantly related to slope aspect in the PPA was RCL, though it was usually higher for trees on south-facing slopes than for trees of the same age on north-facing slopes (Fig. 4d).
Fig. 4 - Growth parameters (mean + standard deviation), of pines on north-facing slopes (black bars) and south-facing slopes (gray bars) in the pine-planted area. (H): total height; (D): diameter at 5 cm above ground level; (H/D): slenderness; (RCL): relative crown length. The numbers above bars are P-values obtained by the Mann-Whitney U test, for the differences between pines of the same age on north- and south-facing slopes.
As shown in Tab. 4, in the NRA pines on the north-facing slopes were significantly older (13.0 ± 4.8 vs. 11.0 ± 4.4 years), taller (2.79 ± 1.43 vs. 2.09 ± 1.36 m), and thicker (diameter: 11.9 ± 7.6 vs. 8.9 ± 6.8 cm) than on south-facing slopes, in stark contrast to relationships observed in the PPA. The tree density was also higher on the north-facing slopes (58.0 ± 91.7 vs. 14.7 ± 40.3 pines ha-1). In the NRA no significant relationship was detected between slope aspect and either slenderness or RCL (Tab. 4).
Tab. 4 - Characteristics of pines growing on slopes with northerly and southerly aspects in the natural regeneration area. (H): total height; (D): diameter at 5 cm above ground level; (H/D): slenderness; (RCL): relative crown length. Values are means ± standard deviation. P-values were obtained after the Mann-Whitney U test.
| Variable | Slope aspect | P-value | |
|---|---|---|---|
| Northerly | Southerly | ||
| Age (years) | 13.0 ± 4.8 | 11.0 ± 4.4 | 0.002 |
| Tree density in polygons (pines·ha-2) | 58.0 ± 91.7 | 14.7 ± 40.3 | 0.01 |
| H (m) | 2.79 ± 1.43 | 2.09 ± 1.36 | < 0.001 |
| D (cm) | 11.9 ± 7.6 | 8.9 ± 6.8 | 0.003 |
| H/D | 0.28 ± 0.11 | 0.29 ± 0.11 | 0.49 |
| RCL (%) | 93.6 ± 4.4 | 94.1 ± 4.1 | 0.84 |
Properties of understorey vegetation in the two woodland types
The understorey vegetation species composition was clearly affected by both woodland type and slope aspect. The first axis of the PCA explains 53.6% of the total observed variance in species composition and clearly separates the two woodland types (Fig. 5), while the second axis clearly separates vegetation on north-facing and south-facing slopes in the NRA, and there is a very weak evidence of separation of the vegetation on north-facing and south-facing slopes in the PPA (Fig. 5).
Fig. 5 - PCA ordination plot showing the distribution of plots in the natural regeneration area (NRA) and pine-planted area (PPA), according to species composition of the understorey vegetation. Species abbreviations: Agr.cap - Agrostis capillaris, Bet.pen - Betula pendula, Cal.epi - Calamagrostis epigejos, Cal.vul - Calluna vulgaris, Car.are - Carex arenaria, Car.eri - Carex ericetorum, Con.maj - Convallaria majalis, Cor.can - Corynephorus canescens, Des.flex - Deschampsia flexuosa, Dry.car - Dryopteris carthusiana, Fag.sil - Fagus sylvatica, Fes.ovi - Festuca ovina, Gal.ver - Galium verum, Hie.pil - Hieracium pilosella, Hyp.per - Hypericum perforatum, Peu.ore - Peucedanum oreoselinum, Pol.odo - Polygonatum odoratum, Pot.rep - Potentilla reptans, Pru.ser - Prunus serotina, Pyr.com - Pyrus communis, Que.rob - Quercus robur, Rub.pli - Rubus plicatus, Rum.ace - Rumex acetosella, Sen.vis - Senecio viscosus, Sol.vir - Solidago virgaurea, Sor.auc - Sorbus aucuparia, Thy.ser - Thymus serpyllum, Vac.myr - Vaccinium myrtillus, Vac.vit - Vaccinium vitis-idaea, Vin.hir - Vincetoxicum hirundinaria.
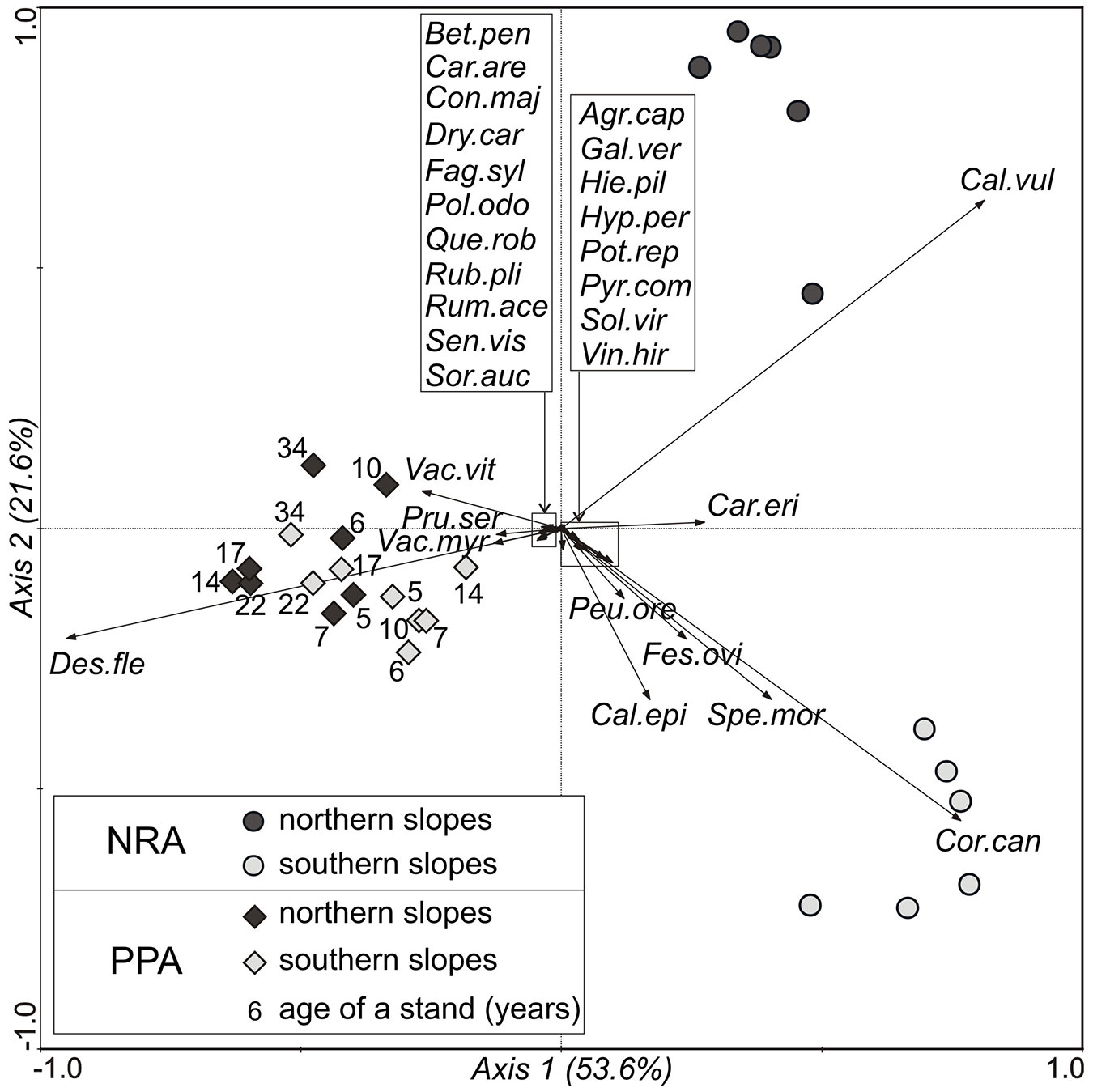
In total 30 vascular plant species were found in the understorey vegetation of the investigated plots. Certain species were clearly abundant in a specific woodland type and/or on slopes with one of the aspects, as shown in Fig. 5. Deschampsia flexuosa dominated on both the north- and south-facing slopes in PPA stands, but neither of these sets of slopes in the NRA. Furthermore, aspect clearly affected the species composition of understorey vegetation in NRA plots, notably Calluna vulgaris dominated on the north-facing slopes, while Corynephorus canescens dominated on south-facing slopes.
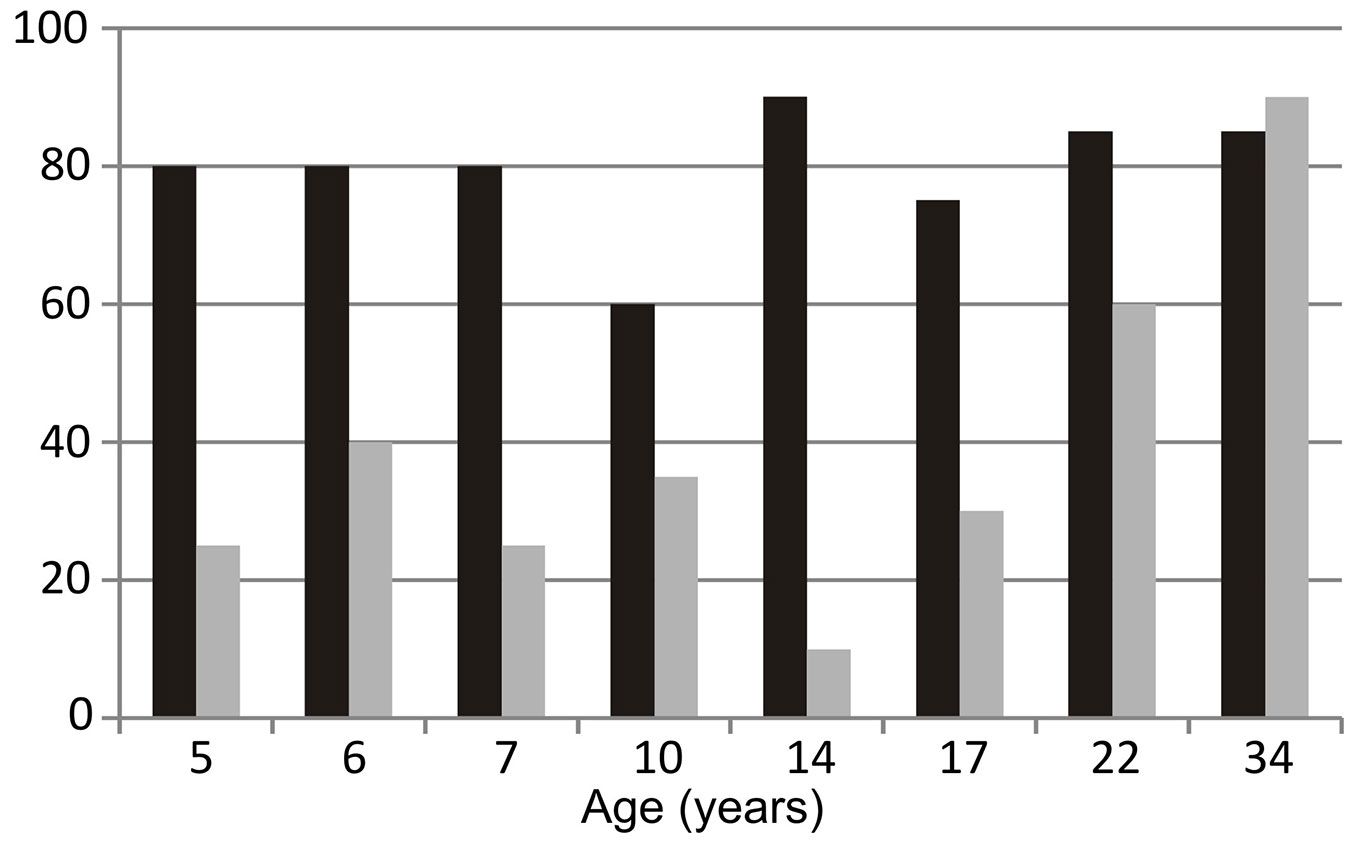
Aspect significantly affected herb cover in the PPA, where it was twice as high on north-facing slopes than on south-facing slopes overall (79 ± 9% vs. 39 ± 25%, respectively; P < 0.01), and higher on north-facing slopes in all but the oldest stand (Fig. 6). In contrast, in the NRA, although aspect clearly affected species composition, it had no effect on herb cover, which was almost identical on north-facing and south-facing slopes (77.5 ± 12.9% and 78.3 ± 7.5%, respectively; P = 0.94).
Fig. 6 - Herb cover, in percentages, on north-facing slopes (black bars) and south-facing (gray bars) slopes in stands of the pine-planted area.
Clear relationships between the calculated Ellenberg’s indicator values and both topographical positions and woodland types were detected (Tab. 5). As expected, the light index (L) values were higher for NRA plots than for PPA plots, in accordance with the more open canopy in the NRA. Moreover, in both woodland types the index was higher for south- than for north-facing slopes, but this distinction was clearer in the PPA. In both woodland types, the moisture index (F) values were higher and the two indices relevant to soil fertility (reaction - R, and nitrogen - N) were lower for north- than for south-facing slopes. However, no consistent differences between slope aspects in temperature (T) or continentality (K) indices were detected in either of the two woodland types (Tab. 5).
Tab. 5 - Ellenberg’s indicator values (means ± standard deviation) for plots on northern and southern slopes in the natural regeneration area (NRA) and the pine-planted area (PPA) for the ecological variables light (L), temperature (T), continentality (K), moisture (F), reaction (R) and nitrogen (N). P-values were obtained after Kruskal-Wallis tests.
| Revegetation type variant |
Slope aspect |
L | T | K | F | R | N |
|---|---|---|---|---|---|---|---|
| NRA | Northerly | 7.3 ± 0.3 | 5.1 ± 0.2 | 3.8 ± 0.2 | 3.8 ± 0.4 | 1.3 ± 0.4 | 1.4 ± 0.4 |
| Southerly | 7.4 ± 0.2 | 5.5 ± 0.1 | 4.5 ± 0.3 | 2.9 ± 0.1 | 3.2 ± 0.4 | 2.3 ± 0.3 | |
| PPA | Northerly | 5.9 ± 0.2 | 5.6 ± 0.4 | 3.4 ± 0.4 | 4.0 ± 0.3 | 2.2 ± 0.2 | 2.7 ± 0.4 |
| Southerly | 6.3 ± 0.3 | 5.4 ± 0.2 | 3.2 ± 0.9 | 3.5 ± 0.4 | 2.4 ± 0.4 | 3.3 ± 0.5 | |
| P-value | - | <0.001 | 0.02 | 0.002 | 0.003 | <0.001 | <0.001 |
Discussion
The growth dynamics of Scots pine stands strongly depends on the space available to the trees in their immediate environments ([21]). The detected differences in pine growth parameters between the two studied woodland types were correlated with variations in tree density at the examined plots. The canopy of a planted pine stand will close some years after planting, but before closure the juvenile trees will have similar space to seedlings in a NRA. This explains the similarities in dimensions of the youngest pines of corresponding ages in the two woodland types. However, in the older PPA stands the canopy had closed and trees had competed more intensely for light, as reflected in their relatively high H and H/D values. Furthermore, such competition is stronger on north-facing slopes than on south-facing slopes, as the former receive less light, and thus the slenderness ratios (H/D) are even higher. In contrast, the canopy appears to remain open longer during the development of stands in the NRA (Fig. 2), thus intraspecific competition for light does not affect pine trees’ growth so strongly. Similar patterns have been observed in a comparative study on Scots pine forests growing at sites with dry, sandy soils in China ([49]).
The NRA results suggest that north-facing dune slopes can offer favorable conditions for forest succession in Central Europe. Similarly, Calvo et al. ([7]) found that the density and height of Pinus pinaster seedlings were higher on north-facing slopes than on south-facing slopes in the three years following a forest fire in north-west Spain. The generally higher moisture and/ or fertility of soils on north-facing slopes could explain these results ([32], [7], [39]). The Ellenberg’s indicator values obtained in this study confirmed that soil moisture was higher on the north-facing slopes of the examined sites. However, as the calculated indices for soil fertility (R and N) were higher on south-facing slopes, the results suggest that soil moisture may influence tree succession on inland dunes such as those at Torun more strongly than soil fertility.
Due to favorable soil conditions, north-facing slopes have also been regarded as superior to south-facing slopes for timber production ([27], [16], [40]). Thus, a surprising finding was that trees growing on north-facing slopes of even-aged PPA stands had lower mean heights and diameters than those growing on south-facing slopes. This may have been due to two factors. First, the north-facing slopes of the studied PPA stands had a higher herb cover (largely of D. flexuosa) than south-facing slopes. This would have affected pine growth in the first years after planting because grasses fiercely compete with young trees for resources ([2], [17], [31], [29]). A similar relationship between grass cover and the growth dynamics of young pines on north- and south-facing slopes of inland dunes in Poland has been previously reported ([35]). The other likely factor is access to light, as Scots pine is a photophilous species ([15]), and the species’ growth on southern slopes may be boosted by their greater insolation, which is positively correlated to photosynthesis rates in P. sylvestris shoots ([28]). Accordingly, Sewerniak et al. ([34]) found that an old-growth pine stand on a south-facing dune slope had larger dimensions than a similarly aged stand on a north-facing slope at the Torun artillery training ground.
The low density of pines on south-facing slopes of the NRA can be explained by the predominance in the understorey vegetation of grasses, which prolong forest succession due to their strong competitive impact in early stages ([31], [11]). Furthermore, competition with herbaceous vegetation is particularly intense in environments with low water availability ([17]), such as south-facing slopes of inland dunes, where moisture contents of the topsoil may be several-fold lower than on north-facing slopes ([34], [35], [36]). Water availability has also been widely identified as a common factor affecting plant succession on dunes ([14], [42]), and the survival, growth and development of managed forests on dry, sandy soils ([50]).
The tree density in the NRA was lower than expected 20 years after a fire, as fires reportedly stimulate the natural regeneration of trees ([19], [23], [3]). Furthermore, significantly higher densities of Scots pine seedlings have been recorded in other post-fire areas in Poland ([12], [33]). However, this study was conducted in a region of relatively low precipitation compared to Central European areas, and the sandy soils of the examined dune ecosystem have poor ability to retain moisture ([20], [34]). These factors hinder seedling survival, especially during dry years when seedlings must compete with weeds for meager rainfall. The scarcity of sources of propagules near the study site could also have contributed to the low tree density in the NRA ([37]). Furthermore, if the residence times of the last fires had exceeded 10 minutes in any parts of the studied plots and generated temperatures exceeding 90 °C, the germinative capacity of the Scots pine seeds in seed banks could have been harmed and the competitor species favored ([26]).
Conclusions
This comparative study extends understanding of the early dynamics of Scots pine stands in inland dune woodlands, and the effects of both slope aspect and standard management. The naturally regenerated and pine-planted stands differed in both species composition of understorey vegetation and pine growth parameters, probably due to large differences in light conditions. Although inland dunes have only slight relief, the aspect was still found to influence vegetation development. The results show that secondary succession of trees proceeds more rapidly on north-facing slopes than on south-facing slopes in inland dune areas of Central Europe. Thus, the south-facing slopes would require more careful attention during attempts to fix inland dunes by establishing or restoring vegetation.
Foresters commonly view inland dunes as uniform sites for forest management, but this study has demonstrated that aspect clearly affects factors that significantly influence tree survival, growth and development on these landforms. Notably, in the pine-planted sites young pines on south-facing slopes had larger mean dimensions than those of corresponding ages on north-facing slopes. This could be due to both the favorable light conditions and lower average herb cover observed on south-facing slopes. The north-facing slopes of young pine plantations had higher weed cover, which negatively affects early Scots pine growth dynamics, particularly in regions with low precipitation. On the dry, sandy dunes of Central Europe, even the relatively low grass species Deschampsia flexuosa can hinder the early growth of Scots pine. The findings, particularly the importance of considering effects of aspect in supposedly homogeneous sites, may facilitate efficient forest management in such environments.
Acknowledgements
This work was supported by the Polish Ministry of Science and Higher Education under grant no. N N305 304840. I am grateful to the administrators of the Torun artillery training ground, both foresters and soldiers, who enabled this research to be conducted in an area that is usually closed to civilians. I would also like to thank Lukasz Mendyk for his help with the GIS analyses. The author is very grateful to two anonymous reviewers for their valuable comments and suggestions on the previous version of the manuscript.
References
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Department of Soil Science and Landscape Management, Nicolaus Copernicus University, Lwowska 1, 87-100 Torun (Poland)
Corresponding author
Paper Info
Citation
Sewerniak P (2016). Differences in early dynamics and effects of slope aspect between naturally regenerated and planted Pinus sylvestris woodland on inland dunes in Poland. iForest 9: 875-882. - doi: 10.3832/ifor1728-009
Academic Editor
Paola Mairota
Paper history
Received: Jun 02, 2015
Accepted: Mar 01, 2016
First online: Jun 17, 2016
Publication Date: Dec 14, 2016
Publication Time: 3.60 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2016
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 53964
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 44661
Abstract Page Views: 3751
PDF Downloads: 4064
Citation/Reference Downloads: 51
XML Downloads: 1437
Web Metrics
Days since publication: 3621
Overall contacts: 53964
Avg. contacts per week: 104.32
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2016): 12
Average cites per year: 1.20
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
The growth dynamics of East European Scots pine (Pinus sylvestris L.) populations - a Lithuanian field trial
vol. 17, pp. 59-68 (online: 06 March 2024)
Research Articles
Amount and distribution of coarse woody debris in pine ecosystems of north-western Spain, Russia and the United States
vol. 7, pp. 53-60 (online: 28 October 2013)
Research Articles
Growth patterns of Scots pine (Pinus sylvestris L.) under the current regional pollution load in Lithuania
vol. 8, pp. 509-516 (online: 12 November 2014)
Research Articles
Coping with spring frost-effects on polyamine metabolism of Scots pine seedlings
vol. 10, pp. 227-236 (online: 27 January 2017)
Research Articles
Twenty years of conversion: from Scots pine plantations to oak dominated multifunctional forests
vol. 10, pp. 75-82 (online: 19 October 2016)
Research Articles
Allometric equations to assess biomass, carbon and nitrogen content of black pine and red pine trees in southern Korea
vol. 10, pp. 483-490 (online: 12 April 2017)
Research Articles
Growing season water balance of an inner alpine Scots pine (Pinus sylvestris L.) forest
vol. 11, pp. 469-475 (online: 02 July 2018)
Research Articles
Scots pine’s capacity to adapt to climate change in hemi-boreal forests in relation to dominating tree increment and site condition
vol. 14, pp. 473-482 (online: 18 October 2021)
Research Articles
A comparison of models for quantifying growth and standing carbon in UK Scots pine forests
vol. 8, pp. 596-605 (online: 02 February 2015)
Review Papers
Biomass, radial growth and regeneration capacity of Aleppo pine, and its possible use as rootstock in arid and degraded areas
vol. 15, pp. 213-219 (online: 16 June 2022)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







