Frost damage to eucalypts in a short-rotation forestry trial in Cumbria (England)
iForest - Biogeosciences and Forestry, Volume 7, Issue 3, Pages 156-161 (2014)
doi: https://doi.org/10.3832/ifor1161-007
Published: Jan 23, 2014 - Copyright © 2014 SISEF
Research Articles
Abstract
Cold is the main climatic constraint to planting eucalypts in Britain and the winter of 2009-2010, the coldest in thirty years, proved particularly challenging for their survival. Damage to transplants planted in May 2009 of two species of eucalypts, Eucalyptus gunnii and Eucalyptus nitens, were assessed over the winter of 2009/2010 at a trial in Cumbria, northern England. Larger trees were found to have exhibited less cold damage by the end of January 2010, but by May there were no significant differences in survival due to tree size. By late January, there were statistically significant differences in damage between E. gunnii and E. nitens with the former being more cold tolerant. However, damage at the end of January, after minimum temperatures of -14 oC, did not appear serious, yet by May the survival of E. gunnii was 37% and for E. nitens was less than 1%. As larger trees exhibited relatively less frost damage it is recommended that intensive silviculture be practiced to ensure trees are between 1 and 1.5 m tall prior to their first winter to reduce the extent of damage through frost.
Keywords
Introduction
Short rotation forestry (SRF) involves growing trees in plantation at a spacing that allows rapid site capture and which are then harvested at a dbh of between 10 and 20 cm ([15]). The wood produced is normally used to substitute fossil fuels as a source of energy. A number of hardwoods were identified as having potential for SRF in the UK ([15]), but the Read Report ([32]) highlighted the potential of eucalypts in sequestration of atmospheric carbon, due to their rapid growth. Of these, two species were identified as having particular potential for the UK: Eucalyptus gunnii Hook.f. and Eucalyptus nitens (Deane and Maiden) Maiden ([15]). There are limited data on growth, but increments of between 3 m3 ha-1 y-1 ([17]) and 18 m3 ha-1 y-1 ([10]) have been reported for E. gunnii at a 7 and 25 year rotation, respectively, and above 30 m3 ha-1 y-1 on a 8 year rotation for E. nitens ([31]).
It is cold that presents the greatest limitation to growing eucalypts in the UK ([19]). Low temperatures have two main negative impacts on the photosynthesis of eucalypts. The first is damage to tissues due to rupture of cells, while the second is photoinhibition of photosynthesis ([11]). Photoinhibition involves a decrease in the efficiency of photosystem II through the combination of cold temperatures and high levels of sunlight ([6]). Photoinhibition occurs least and recovery is most rapid in the most cold-tolerant eucalypts ([16]).
Furthermore, low soil temperatures are known to decrease absorption of water by roots (Teskey et al. 1984, in [8]). When the soil is frozen uptake can be seriously disrupted; generally, soil temperatures of -1 °C or less will prevent water uptake and can increase dehydration (Larcher 1957, in [4]). During periods of warm air temperatures with frozen ground, trees must rely on moisture stored in sap reserves and smaller trees will deplete these reserves faster ([4]).
Eucalypts have four main ways of producing leafy shoots; naked buds in leaf axils, accessory buds, dormant (epicormic) buds and buds in lignotubers. The latter two, dormant and lignotuber buds are particularly important in producing shoots after significant damage, such as fire or frost. However, it is the naked buds and accessory buds that normally contribute to crown development. The naked buds primarily contribute to the development of leafy shoots, with accessory buds providing an alternative if the naked buds are damaged (Jacobs 1955, in [9]). The naked buds grow when temperatures are above a certain minimum, enabling potentially high productivity, especially when grown as exotics ([2]) as this strategy allows growth through much of the year. Unlike most temperate trees, photoperiod has no effect on growth ([23]). Davidson et al. ([11]) note that maximum winter growth rates for E. nitens (Deane and Maiden) Maiden in a plantation in Tasmania were only slightly less than maximum rates in summer. However, this lack of dormancy also leaves eucalypts vulnerable to damage through chilling ([11]). The cold winters experienced in the UK, relative to those of Australia, means that only a limited range of species (those that are from sub-alpine areas of Australia) have survived.
Hardening is a process crucial to providing resistance to cold and also speeds up the recovery time of photsosynthesis, following a period of cold ([11]). In eucalypts a progressive decline in temperature enables hardening within just a few days ([30], [23]). Harwood (1980, in [18]) describes the importance of hardening, noting that there is little difference in frost resistance between sub-alpine species of eucalypts when they are in an unhardened state, yet when hardened they exhibit considerable variation. In sub-alpine eucalypts hardening is initiated through low temperatures, rather than reduced photoperiod (Eldridge 1969, in [1]); the crucial temperature for initiating hardening being between 2 °C ([23]) and 4 °C ([12]). However, both the level and duration of cold is important to the hardening process and it is a characteristic only of those eucalypts from colder climates ([33]).
Hardening does not seem to increase markedly the ability of cold-tolerant eucalypts to limit damage through tolerating supercooling of their tissues, rather it seems to confer resistance to cold through other means ([33]). The mechanism involves an increase in concentration of soluble sugars, stabilising cell membranes and possibly also providing photosynthetic precursors enabling more efficient winter photosynthesis ([1]). Another chemical associated with cold hardiness is anthocyanin, a pigment which is thought to act through reducing absorption of light during photoinhibition and also possibly through a role of quenching antioxidants ([7]). In frost resistant eucalypts, damage occurs at temperatures well below that which ice forms in the tissues, and so death of cells appears to be related to dehydration (Olien 1978, in [36], Steponkus 1984, in [36]). As the water potential of ice is lower than liquid water, freezing draws water from the cells and causes them to dehydrate. If this loss of water is sufficient it can cause disruption to cell membranes resulting in leakage ([24]). This injury, caused by frost dehydration occurs in hardened individuals at a lower temperature than unhardened ones ([36]) and so unseasonal cold is particularly damaging. In a comparison of eucalypt species, the cold-resistant E. gunnii was found to respond rapidly to lower temperatures enabling it to cope with the development of extracellular ice and the associated dehydration of tissues. Scarascia-Mugnozza et al. ([33]) and Valentini et al. ([36]) also noted that the capacity of cold tolerant eucalypts to retain intracellular water was considerably increased by cold hardening.
Recent work undertaken in Ireland has focused on two important aspects of cold tolerance in eight species of eucalypt: lethal temperature and the pattern of hardening (Black, unpublished data). Investigation of LT50 (lethal temperature for 50% of the shoots) showed considerable variation between species. Results also showed that the ranking of species in terms of those most cold tolerant differed between winter 2010-2011 and winter 2011-2012. These differences were probably due to different patterns of hardening in the two winters, the earlier winter being colder than the later one. Further investigation showed that the rate of hardening varied between eucalypt species. Black (unpublished data) suggests that when selecting species, LT50 and the rate of hardening should be combined to create a measure of cold tolerance.
A polar air mass moving from continental Europe brought bitterly cold conditions to Great Britain during December 2009 and January 2010, resulting in the coldest winter in England for over thirty years ([20]), specifically since 1978/1979 ([21]). Across the UK, the mean temperature was 2 °C below the 1971-2000 average, with the most severe cold being in the north of the country. For northern Scotland it was the coldest winter on record and for England the ninth coldest since 1910. For northern England the lowest recorded temperature was -17.6 °C on 7 January 2010 at Woodford, near Manchester. This was the lowest temperature for that location on record ([29]).
The objectives of this study were to examine whether there were significant differences in frost damage and survival between E. gunnii and E. nitens and between larger and smaller trees over the extreme winter period of 2009/2010.
Methods
The methods section is divided in three parts, the first describing the characteristics of the trial, the second the approach used to collect data and the third the methods used to analyse frost damage and survival.
Description of the trial
A trial adopting a randomised complete- block design was established close to the Newton Rigg Campus of the University of Cumbria (54° 40’ 56” N, 2° 47’ 22” W), testing five tree species in six replicates. The species used had been identified in Hardcastle ([15]) as being hardwoods with sufficiently rapid growth to be used in SRF, to produce biomass for energy. These comprised, sycamore (Acer pseudoplatanus L.), alder (Alnus glutinosa L.), ash (Fraxinus excelsior L.) and two eucalypts; Eucalyptus gunnii and Eucalyptus nitens. Container-grown seedlings were planted using a “T” notch and established in 60cm tubes. The area was stock-fenced. Native and naturalised species were planted in November 2008, while the eucalypts were planted later, in late April 2009, with the aim of avoiding late spring frosts.
The trial was originally under grass pasture and the soil was a clay loam brown earth and slightly acid (pH 6). Bulk density was 0.76 at 0 cm to 15 cm depth and 1.07 at 15 cm to 30 cm depth. Soil nitrogen was 5.45 tonnes ha-1 and 4.35 tonnes ha-1 at 0 cm to 15 cm and 15 cm to 30 cm depth respectively ([5]). A complete-weeding approach was adopted to kill the pasture grasses using a combination of propazymide and glyphosate. Survival of trees in November, at the end of the first growing season, was over 87% for all species, E. gunnii having 87% and E. nitens having 96% survival. Growth was rapid, E. nitens attaining a mean height of 1.09 m and E. gunnii of 1.39 m, a reflection of a rigorous complete weeding regime and an unusually wet and warm summer ([22]). However this rapid growth led to instability in some of the larger trees, reflecting an inbalance between root development and shoot growth.
The origin of the eucalypts was given as E. nitens “Central Victoria” and of E. gunnii as being Region 94, which is Australia (Watson, pers. comm.). The E. gunnii supplied contained hybrids of E. gunnii and E. nitens and the foliage of the E. gunnii was not glaucous. In the opinion of Purse (pers. comm) because of these two factors, the seed was probably from the only commercial source of E. gunnii, a stand in Dipton, New Zealand. Purse (pers. comm.) also attributed the E. nitens to being central Victoria origin but Dipton, New Zealand provenance.
Methods for data collection
The trial at Newton Rigg followed a randomised complete block design with six replicates and plots containing 80 trees each. For this survey a sub-plot of 24 trees was created in each of these plots. All 24 trees within this sub-plot were scored on an eleven point system for frost damage on the 31 January 2010, and then again on the 1 May 2010. The system for scoring frost damage was based on one used by Evans ([13]), except that three parts of the trees were scored separately: the lower stem (within the tree shelters), the upper stem (outside the tree shelters), and the foliage, whereas Evans ([13]) scored crown foliage only. The scoring ranges from 0, which is no visible damage, to 10 which represents 91-100% damage. As a measure of necrosis, the extent to which the cambium had been blackened and the extent of discolouration of the foliage (from healthy green to damaged khaki) was used.
The trial was originally established to test yield of potential SRF species and so maintaining the plots at full stocking was important. In April 2010 the trial was beaten-up and as it was not clear whether the trees would recover, the decision was made to beat up half of the trees in the plots. As such, the size of the initial frost damage plots was reduced from 24 to 12 trees as the other half of the plot had been replaced with new trees. For E. nitens it was clear the trees would not recover and so all trees were replaced. For E. gunnii, 37% of the original trees were alive and these were not replaced.
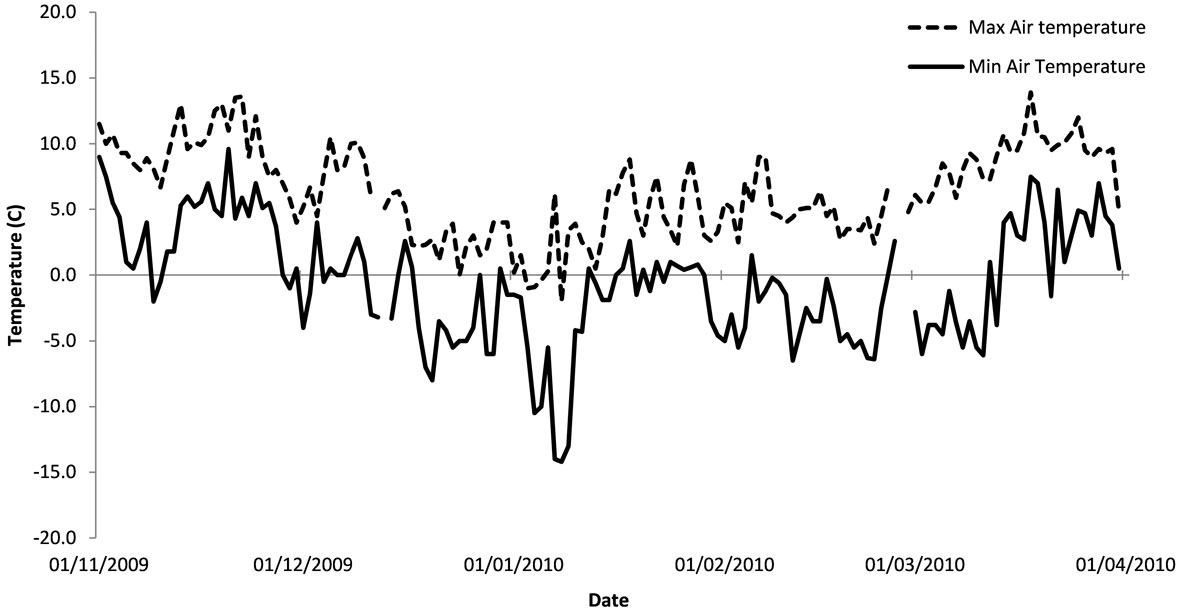
The climatic records for the winter of 2009/2010 were obtained from the weather station 1 km away at the Newton Rigg Campus of the University of Cumbria. Fig. 1 shows the daily minimum and maximum air temperatures over the period between November 2009 and March 2010 at Newton Rigg Campus. In addition to the extreme cold, the daily range in temperature was considerable, reaching 20 °C during one twenty four hour period, varying from -14 to 6 °C. Twenty four hour temperature fluctuations of nearly 10 °C were frequent during the winter, due to the combination of cold nights and clear sunny days. During December, January, February and March, the grass minimum temperature fell below freezing on 24, 28, 25, 21 and 14 days respectively. On the 9 January 2010 the grass minimum dropped to a low of -17 °C. The period of sunshine was generally above the 1971-2000 average ([21]). During the period between mid December and mid January there was almost constant snow cover, with depths of up to 19 cm. During the remainder of the winter there were only infrequent, small falls of snow, the ground being bare for much of the time.
Fig. 1 - Maximum and minimum air temperatures over the winter of 2009/2010 using data from the Newton Rigg weather station.
Analysis of data
The distribution of frost damage score data for cold damage at lower stem, upper stem and foliage by species was tested for normality before and after arcsine transformation. Significant departure from normal distribution was found (p<0.0001), so a non parametric Kolmogorov-Smirnoff test was used to test for differences in the frost damage scores between the two species.
The role of tree size on cold damage was investigated by dividing the trees into quartiles by their height in January 2010. For these quartiles, the data on cold damage and survival was tested for conformance to a normal distribution. By quartile and by block scores for cold damage and arcsine cold damage for E. gunnii and for E. nitens were found to be significantly different from normal; therefore non-parametric Kruskal-Wallis tests and Mann-Whitney U tests were applied to the data.
A Chi-squared test was used to determine whether significant differences exist in survival between the two eucalypts, between the quartiles by height and between blocks.
Results
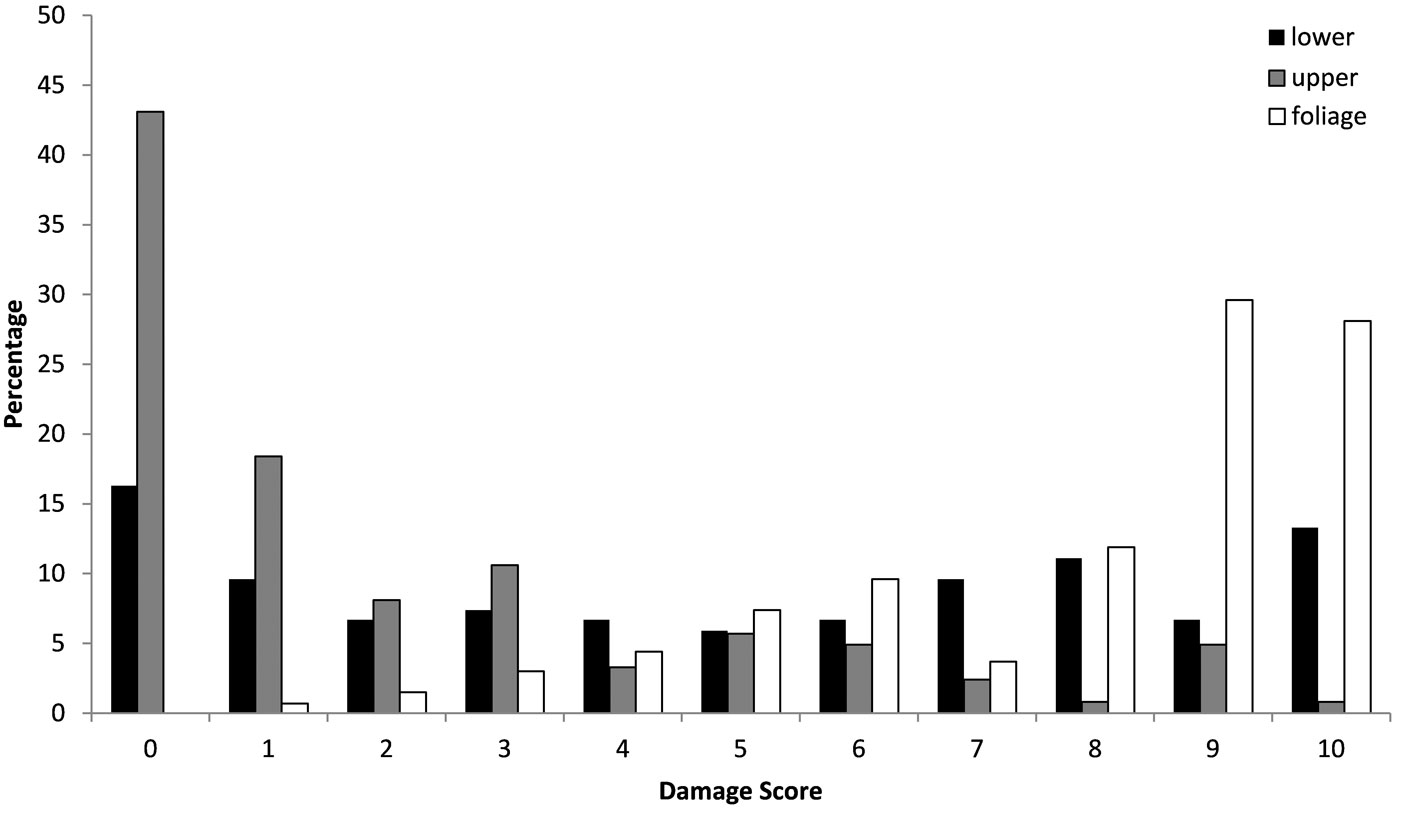
The pattern of frost damage noted on 31 January 2010 in the lower stem, upper stem and the foliage was compared between E. gunnii and E. nitens using a Kolmogorov- Smirnoff test, finding significant differences (P<0.001) between the two species. E. nitens was found to be more susceptible to damage by frost to stem and foliage, as proved from higher scores for frost damage. Despite very low temperatures in January of -14 °C minimum air temperature or -17 °C grass minimum, the E. gunnii showed relatively little visible damage (Fig. 2), whereas the damage to E. nitens was very obvious, particularly to its foliage (Fig. 3). In January, survival remained high with that of E. gunnii being 93% and of E. nitens being 93%. The results of the assessment of 1 May 2010 showed that there had been a substantial increase in damage, particularly to foliage of both species and considerable further mortality. Survival of E. gunnii had declined to 35%, whereas for E. nitens it had dropped to less than 1%.
Fig. 2 - Frost damage in upper stem, lower stem and foliage of E. gunnii. (0): no damage; (1): 1-10% damage; (2): 11-20% damage; (3): 21-30% damage; (4): 31-40% damage; (5): 41-50% damage; (6): 51-60% damage; (7): 61-70% damage; (8): 71-80% damage; (9): 81-90% damage; (10): 91-100% damage.
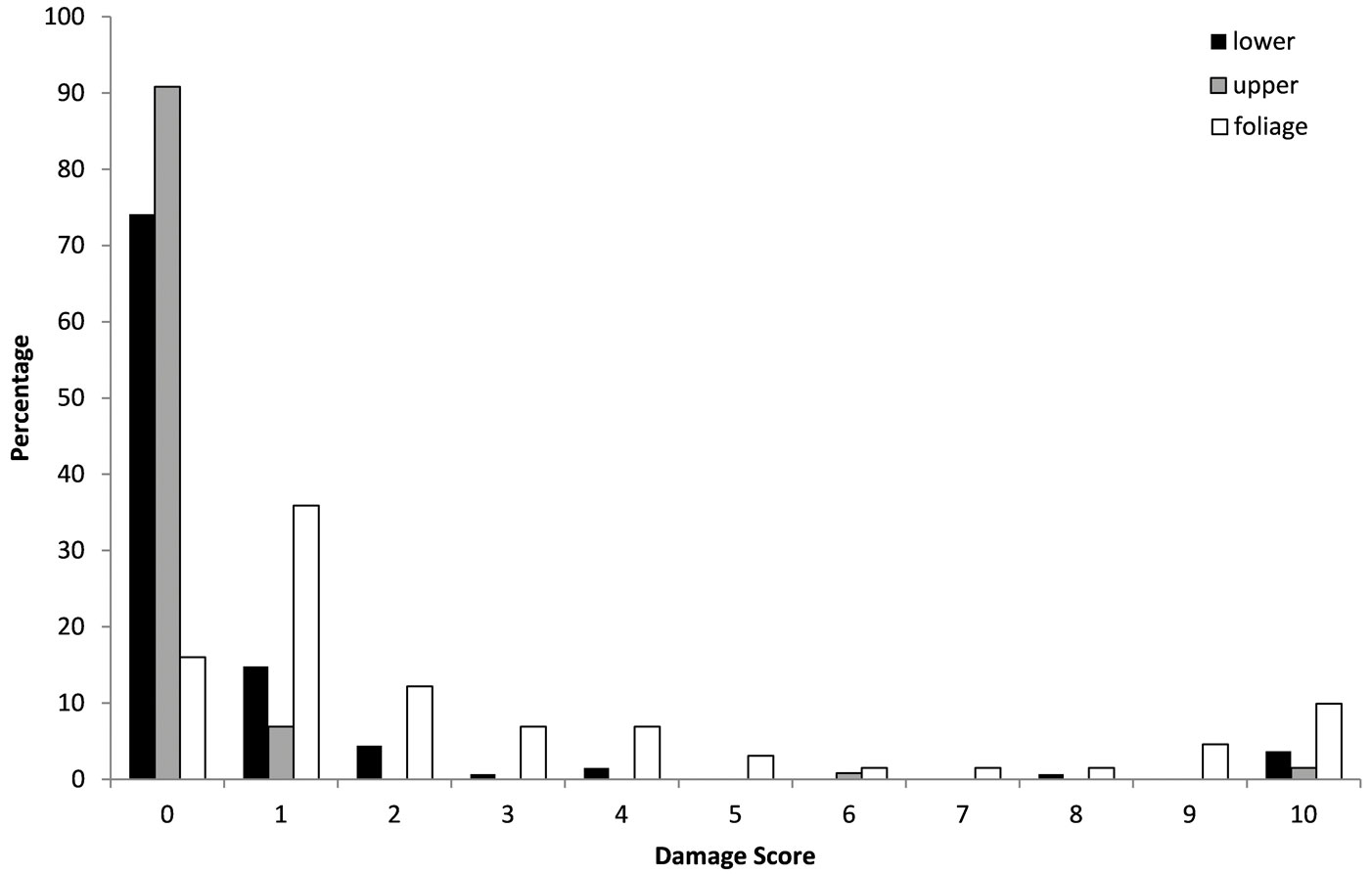
Fig. 3 - Frost damage in upper stem, lower stem and foliage of E. nitens. (0): no damage; (1): 1-10% damage; (2): 11-20% damage; (3): 21-30% damage; (4): 31-40% damage; (5): 41-50% damage; (6): 51-60% damage; (7): 61-70% damage; (8): 71-80% damage; (9): 81-90% damage; (10): 91-100% damage.
There appeared to be a relationship between tree height and damage. The median heights for each quartile are shown in Tab. 1 with the overall ranking of damage by quartile. In E. gunnii damage to lower stem, upper stem and foliage was greatest in the quartiles containing the smaller trees. A Kruskal-Wallis test was used to examine differences in damage between quartiles. There were significant differences between quartiles for damage in the lower stem (p=0.023), upper stem (p=0.025) and foliage (p=0.011). Mann-Whitney U tests were used to identify where these differences originated. These are described in Tab. 2 and the damage to the upper stem and foliage for the quartile with the smallest trees was significantly different from the other quartiles. There were other significantly significant differences but the pattern was not clear (Tab. 2).
Tab. 1 - Ranking of damage score in January 2010 by tree height, divided into quartiles, where (1): lowest damage to (4): highest damage.
| Height of trees by Quartile |
E. gunnii | E. nitens | ||||||
|---|---|---|---|---|---|---|---|---|
| Median height (cm) | Damage Score | Median height (cm) | Damage score | |||||
| Lower stem | Upper stem | Foliage | Lower stem | Upper stem | Foliage | |||
| Smallest | 98 | 3 | 4 | 4 | 73 | 4 | 4 | 4 |
| Small | 131 | 4 | 3 | 3 | 108 | 2 | 2 | 2 |
| Large | 155 | 1 | 2 | 2 | 122 | 3 | 1 | 3 |
| Largest | 175 | 2 | 1 | 1 | 147 | 1 | 3 | 1 |
Tab. 2 - Probabilities from Mann-Whitney U tests comparing damage in E. gunnii between quartiles of tree height. (SL): stem low; (SH): stem high; (FO): Foliage.
| Prob | Quartile | |||
|---|---|---|---|---|
| Small | Large | Largest | ||
| Quartile | Smallest | SL: 0.960 | SL: 0.012 | SL: 0.313 |
| SH: 0.046 | SH: 0.042 | SH: 0.022 | ||
| FO: 0.041 | FO: 0.014 | FO: 0.002 | ||
| Small | - | SL: 0.003 | SL: 0.153 | |
| SH: 0.976 | SH: 0.613 | |||
| FO: 0.578 | FO: 0.194 | |||
| Large | - | - | SL: 0.075 | |
| SH: 0.631 | ||||
| FO: 0.501 | ||||
For E. gunnii an examination was undertaken to determine whether survival in May 2010 after months of freezing conditions was related to tree height. While survival was lowest (30%) in the quartile containing the largest trees it was next lowest in the smallest trees (32%) and highest survival was in the trees in the second smallest quartile (41%). A Chi-squared test indicated that differences between survival in the quartiles were not significant (p=0.802).
As with E. gunnii, the E. nitens trees were divided into quartiles by height in January 2010 and a Kruskall-Wallis test was used to identify if the differences in frost damage by quartile were significant. Differences between quartiles in terms of foliage and lower stem damage were highly significant (p<0.0001). Mann Whitney tests were applied to the foliage and lower stem data by quartile to identify where these differences lay and only the smallest quartile showed damage significantly different to others (Tab. 3).
Tab. 3 - Probabilities from Mann-Whitney U tests comparing lower stem damage and foliage damage in E. nitens between quartiles of tree height.
| Prob | Quartile | |||
|---|---|---|---|---|
| Small | Large | Largest | ||
| Quartile | Smallest | SL: 0.0001 | SL: 0.001 | SL: 0.0001 |
| FO: 0.0001 | FO: 0.0001 | FO: 0.0001 | ||
| Small | - | SL: 0.878 | SL: 0.878 | |
| FO: 0.586 | FO: 0.586 | |||
| Large | - | - | SL: 0.241 | |
| FO: 0.169 | ||||
A Kruskal-Wallis test showed that differences in damage in January between blocks for E. gunnii were not significant for lower stem and foliage, but were highly significant for upper stem. The level of upper stem damage was however low in all blocks. A similar analysis of damage in E. nitens in January and E. gunnii in May showed no significant differences in damage in lower stem, upper stem and foliage by block. There were insufficient E. nitens surviving in May to conduct an analysis of damage by block.
Survival of E. gunnii by block by May varied from 54% to 20% and a Chi-squared test showed the differences not to be significant (p=0.116). Despite evidence of epicormic growth in trees that were left when part of the plots were replanted, none of those that were recorded as being dead recovered during the summer. There were insufficient trees surviving of E. nitens in May to undertake a similar analysis of survival between January and May by quartile or by block.
Discussion
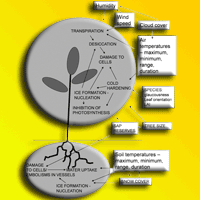
The influences determining the degree of damage to eucalypts from cold are complex and are related to a number of factors which are summarised in Fig. 4. However well- adapted temperate eucalypts are to the UK climate, the winter of 2009/2010 was the coldest in the last thirty years ([20]) and the combination of severe cold and almost three months of days where temperatures dropped below freezing will have caused severe plant stress. Further, due to clear skies, the range in temperature over 24-hour periods was considerable, resulting in variation in temperature of 20 °C during one twenty four hour period in January, during which the trees would have experienced periods of freezing and thawing of above ground and below ground tissues. Two factors may have reduced damage somewhat: the gradual but steady decline in temperatures during December (Fig. 1) would have allowed the trees to harden and also damage may have been mitigated to a degree by the insulating layer of snow that lay on the ground from mid December to mid January, protecting the roots from the extreme air temperatures.
Fig. 4 - Summary of external and internal physiological factors affecting frost damage
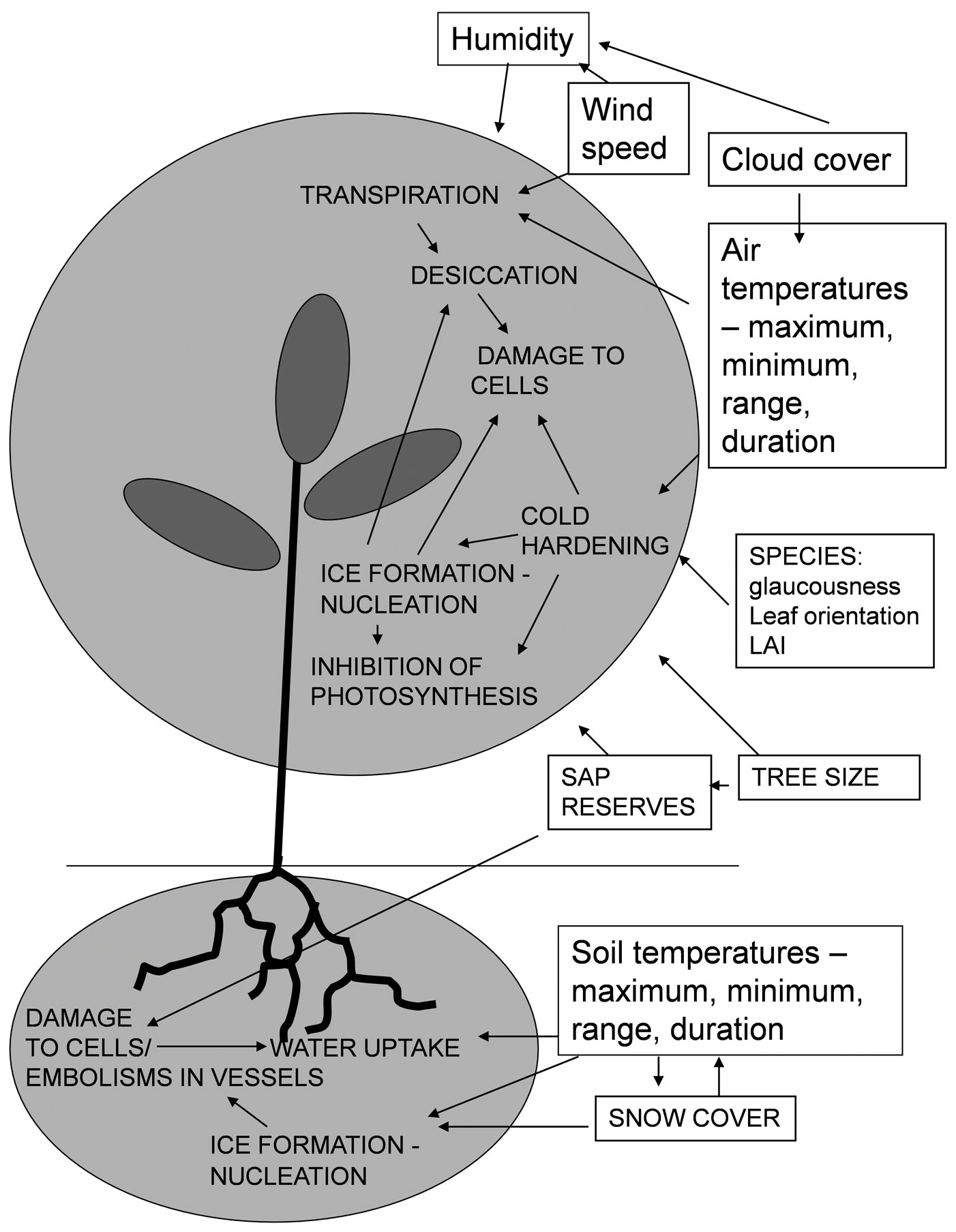
The two species of eucalypt tested in the trial have different climatic tolerances, including their capacity to resist cold ([3]). It is E. gunnii that inhabits a colder alpine environment in Tasmania, compared with the montane, lower latitude areas occupied by E. nitens on the main portion of Australia. E. gunnii is noted as being one of the most cold hardy species, being highly resistant even in an unhardened state ([12]). This is supported by results from this trial; by the end of January, E. nitens had suffered significantly worse damage then E. gunnii from the cold (Fig. 3 and Fig. 4). However, in early February many of the terminal buds of E. nitens still appeared green, flexible and undamaged (Hepburne-Scott. pers. comm.). By May the injury to the trees had increased markedly and only 35% of the E. gunnii remained alive, while less than 2% of the E. nitens had survived.
Work by Black (unpublished data) in Ireland has shown that there are considerable differences between cold-tolerant eucalypts not only in terms of lethal minimum temperatures but also in their pattern of acclimation to cold. Absolute lethal temperature and ranking by seasonal variance in lethal temperature were combined to produce an overall rank of cold tolerance. Of seven species of eucalypts, E. nitens was found to be the poorest in terms of cold tolerance, with E. gunnii being fourth out of the seven species. This contradicts other work that suggests E. gunnii is particularly cold-tolerant ([12]).
The evaluation of cold damage to the trees was undertaken using a visual scoring system, but a more reliable and quantifiable approach for evaluating damage to the foliage would have been to measure chlorophyll fluorescence as detailed in Perks et al. ([25]). Also, the impacts of cold damage are more often measured under controlled conditions, for example using a freezing cabinet. This is because of the many factors that influence cold damage in the field, such as variation in micro-topography and sky exposure, in the frost hardiness of the trees across and between seasons and between and within populations.
The assessment showed clearly that woody tissues suffered less extensively from damage from the cold than the foliage in both species of eucalypt, an observation supported by others ([33]).
In the assessment of damage carried out in January 2010 there was considerable variation in frost injury between trees, even those adjacent to one another. While some individuals exhibited almost complete damage to foliage, others remained almost uninjured. This variation could be due to differences in: the genetic composition of individuals, the size of the individuals, the micro site they occupied or their treatment during planting and tending. Considerable variation in the frost resistance of provenances and individuals within provenances has been noted in both E. gunnii ([26], [27], [28]) and E. nitens ([34], [35], [14]) in their natural habitats. A study of frost tolerance of 101 origins of E. nitens planted in Tasmania showed the western provenances of the central highlands of Victoria and those from New South Wales to be superior ([34]), while from early results Evans ([13]) found origins of E. nitens from Victoria were most cold hardy in trials in Great Britain. For E. gunnii there is convincing evidence from British trials ([13], [10]) that provenances from Lake MacKenzie are more frost tolerant.
The finding that larger trees are more resistant to damage highlights the importance of obtaining rapid early growth so as to obtain a tree of 1-1.5 m height before the onset of winter. Rapid growth is important, as larger trees have greater sap reserves and once trees reach 2-6 m the sensitive growing tips are usually above mild growing season radiation frosts ([12]). Furthermore, larger trees exhibit greater physiologi- cal maturity and tolerance to environmental stresses than smaller trees of the same age. Ensuring rapid establishment is therefore crucial, including effective weed control and ensuring the trees receive adequate nutrition. Furthermore, good nutrition has also been shown to be important through reducing the extent of photoinhibition in seedlings of E. nitens during cold periods ([6]). The increase in damage to the very largest trees at the trial may be explained by an imbalance between the root/shoot ratio; many had proven to be unstable being prone to lean and had required additional staking.
The effects of frost can be difficult to predict as both fast recovery and long-term deleterious effects have been noted (Ball 1994, in [18]). The effects of repeated frosts can have a compounding effect on growth and survival, especially in trees in a phase of rapid, early growth, where death of mature leaves and developing shoots can delay investment of resources into new leaves (Ball et al. 1997, in [18]). The weak growth in the summer of 2010 and complete mortality of the E. gunnii that survived the winter of 2009-2010 in the subsequent, milder winter supports this observation. The results from this trial show that following severe cold damage it is best to replant young eucalypts as recovery is unlikely, and even those individuals that have survived are likely to have lower growth and survival.
Conclusions
The results of this study support the results of others from both field trials ([13]) and laboratory tests ([3]) that E. gunnii is more cold tolerant than E. nitens. Only two of the 144 trees assessed of E. nitens survived by May 2010, compared with 43 of the 144 trees of E. gunnii. Despite the once in thirty year conditions experienced in winter 2009-2010 and the trees being less than one year on the site, the better survival of E. gunnii suggest in terms of adaptability that it is a species that could be used for producing woody biomass even in northern parts of Britain. Also, it is probable that survival of E. gunnii at the site would have been enhanced by use of material with the origin of Lake Mackenzie, that best adapted to British conditions ([13], [10]). Relative damage, but not survival is related to the size of the young trees, with larger trees being more resistant. It is therefore imperative that transplants, through intensive silviculture, provision of adequate nutrition, are given the greatest opportunity to establish effectively and grow rapidly before their first winter, when they are particularly vulnerable.
Acknowledgements
The authors would like to thank the following people: Alastair Chalmers of Pyroclassic Fires for funding half the establishment costs of the trial; Pat Tyers, Graham Leslie and Chris Leslie for helping with data collection and also Kevin Black for providing a draft manuscript of his report on cold tolerance studies on eucalypts in Ireland. They also thank Dr. Conor O’Reilly and Dr. Euan Mason for highlighting areas requiring improvement in the original manuscript.
References
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
National School of Forestry, University of Cumbria, Penrith Campus, Newton Rigg, CA11 0AH Penrith, England (UK)
School of Geosciences, University of Edinburgh, Crew Building, The King’s Buildings, West Mains Road, EH9 3JN Edinburgh, Scotland (UK)
Centre for Forestry and Climate Change, Forest Research Northern Research Station, EH25 9SY Roslin Midlothian, Scotland (UK)
Corresponding author
Paper Info
Citation
Leslie AD, Mencuccini M, Perks M (2014). Frost damage to eucalypts in a short-rotation forestry trial in Cumbria (England). iForest 7: 156-161. - doi: 10.3832/ifor1161-007
Academic Editor
Marco Borghetti
Paper history
Received: Oct 22, 2013
Accepted: Jan 13, 2014
First online: Jan 23, 2014
Publication Date: Jun 02, 2014
Publication Time: 0.33 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2014
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 59838
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 48385
Abstract Page Views: 5364
PDF Downloads: 4502
Citation/Reference Downloads: 21
XML Downloads: 1566
Web Metrics
Days since publication: 4497
Overall contacts: 59838
Avg. contacts per week: 93.14
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2014): 5
Average cites per year: 0.42
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Interaction between planting spacing and wood properties of Eucalyptus clones grown in short rotation
vol. 14, pp. 12-17 (online: 02 January 2021)
Research Articles
Variation of wood and bark density and production in coppiced Eucalyptus globulus trees in a second rotation
vol. 9, pp. 270-275 (online: 08 September 2015)
Research Articles
Impact of rotation length of Eucalyptus globulus Labill. on wood production, kraft pulping, and forest value
vol. 15, pp. 433-443 (online: 20 October 2022)
Research Articles
Validation of models using near-infrared spectroscopy to estimate basic density and chemical composition of Eucalyptus wood
vol. 17, pp. 338-345 (online: 03 November 2024)
Research Articles
Relationship between frost hardiness of adults and seedlings of different tree species
vol. 7, pp. 282-288 (online: 01 April 2014)
Research Articles
Coping with spring frost-effects on polyamine metabolism of Scots pine seedlings
vol. 10, pp. 227-236 (online: 27 January 2017)
Research Articles
NIR-based models for estimating selected physical and chemical wood properties from fast-growing plantations
vol. 15, pp. 372-380 (online: 05 October 2022)
Research Articles
Effect of stand density on longitudinal variation of wood and bark growth in fast-growing Eucalyptus plantations
vol. 12, pp. 527-532 (online: 12 December 2019)
Research Articles
Kinetic, morphophysiological, and biochemical parameters as a strategy to select clones of Eucalyptus spp. more efficient in phosphorus uptake
vol. 18, pp. 212-222 (online: 09 August 2025)
Research Articles
Potential for utilization of wood ash on coastal arenosols with limited buffer capacity in KwaZulu-Natal and its effect on eucalypt stand nutrition and growth
vol. 10, pp. 180-188 (online: 19 November 2016)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords







