Evaluating humus forms variation in an unmanaged mixed beech forest using two different classification methods
iForest - Biogeosciences and Forestry, Volume 5, Issue 5, Pages 272-275 (2012)
doi: https://doi.org/10.3832/ifor0632-005
Published: Oct 24, 2012 - Copyright © 2012 SISEF
Technical Reports
Abstract
Humus is an important part of terrestrial ecosystems and can be considered as an indicator of ecosystem functioning status. Morphologic method is an inexpensive and easy way to study humus forms. This study investigated two morphological methods that have been applied in beech stands of Hyrcanian forest and compared them to assess which one is more appropriate for these ecosystems. Therefore an unmanaged beech stand was selected and 320 humus profiles were considered in it. In each profile the humus form was determined as to suborder level according to two morphological methods. The results showed that the average thickness of organic and organo-mineral horizons (OL, OF, OH and Ah) in the study site was 2, 0.6, 0.3 and 3.6 cm, respectively. Also the two different morphological methods used in the study site had different functions and outputs. According to both methods the Mull order was the dominant humus form in the study site. The number of humus suborders found in the study site was different in the two methods and indicates their different ability in describing humus forms in the study site
Keywords
Humus Classification, Mixed Beech Forests, Hyrcanian Forest, Organic Horizons
Introduction
Decomposition of organic matter plays a vital role in nutrient cycling through which organically bound nutrients mineralize and become available for plant uptake. In infertile soils the role of decomposition processes becomes even more significant in nutrient cycling, since almost all plant available nutrients in these ecosystems originate from plant debris ([16]). Different types of humus forms develop within the top-soil throughout the process of biotransformation of dead organic matter. Climatic conditions, soil characteristics and vegetation types are among the variables that can affect this process ([8]). Humus forms have been considered as one of the principal components of terrestrial ecosystems ([15]). Since humus forms develop at shorter time scales than soil types, they have to be classified independently ([7]). Humus forms are indicators of ecosystem functioning and characteristics, implying that the study of humus forms provides information on site conditions such as availability of moisture, soil acidity, nutrient status ([17]) and decomposition rates ([2]). Humus represents the complex organic substances resulted from the decomposition of plant and animal fragments. These materials are more stable, colloidal, and mostly in connection with the mineral component of the soil. Humus forms commonly are comprised of organic and organic-enriched mineral horizons at the soil surface, and are considered as natural bodies such as the underlying soils ([8]). The morphological study of humus forms has begun since the 19th century ([10]) and until now many morphological methods of classification have been developed worldwide ([3], [8], [6], [17], [19], 2011). The morphological study of humus forms, in comparison to chemical methods, is inexpensive, easier to manage on the field and can easily be instructed to the forest staffs.
Hyrcanian forests, covering about 1.9 million ha, are broad-leaved deciduous forests forming a green belt along southern and south-western coasts of the Caspian Sea. They are unique in genetic variation, biodiversity, commercial productions and many other environmental services ([14], [12], [13], [4]). The first morphological study on humus forms in Hyrcanian forests was done by Sajedi et al. ([15]). They classified the humus forms in a pure beech forest using a Canadian grid by Green et al. ([8]). Their results showed that Moder was the dominant humus form in the studied stands. In second attempt, Waez-Mousavi ([18]) applied a European humus classification method ([19]) to the study of humus forms in some mixed beech stands of Hyrcanian forests. He found that humus forms belonging to the Mull group were dominant in this area. The present study aimed to compare the two morphological methods of classification which have been applied in beech stands of Hyrcanian forest and to assess the most appropriate method for the ecological characterization of these ecosystems.
Materials and methods
Study site
This investigation was carried out in parcel 32 of compartment 1 in Shast Kalate (Bahram Nia) forest, experimental forest of Gorgan University of Agricultural Sciences and Natural Resources, a virgin mixed deciduous forest covering an area of about 3716 ha and located in the north of Iran (36°41´ to 36°45′ N and 54°20′ to 54°24′ E), with an average annual precipitation of about 650 mm, and an altitude ranging from 100 to 1000 m a.s.l. The mentioned parcel, with an area of about 80 ha and an altitude of about 900 m above a.s.l.,is mostly covered with Fagus orientalis Lipsky (oriental beech) mixed with Carpinus betulus L. (common hornbeam) and Parrotia persica (DC.) C.A. Mey (Persian ironwood tree). The aforementioned parcel is a permanent plot for long term studies, established on brown forest soil with mostly sandstone as bedrock.
Data collection
In 2009, 320 humus profiles were randomly examined within old-growth mixed beech-dominated stands in the parcel. In each humus profile humus classification units and subunits were determined according to Canadian ([8]) and European ([19]) methods. Both classifications are based on morphological characteristics of organic (L, F, H or OL, OF and OH, respectively) and organo-mineral (Ah or A, respectively) horizons, such as their presence or absence, thickness, structure, animal or fungal activity, etc., among which horizon thickness and structure were the most important. The thickness was measured as to the nearest mm and the structure was assessed to the naked eye or with a 5-10 X magnifying hand lens.
Results
The average thickness of organic and organo-mineral horizons (OL, OF, OH and A) in the study site was 2, 0.6, 0.3 and 3.6 cm, respectively (Tab. 1).
Tab. 1 - Organic and organo-mineral horizon thickness.
| Parameters | N | Range (cm) | Mean (cm) | SD |
|---|---|---|---|---|
| L=OL | 320 | 0-7.5 | 2.0 | 0.90 |
| F=OF | 320 | 0-3 | 0.6 | 0.57 |
| H=OH | 320 | 0-3.5 | 0.3 | 0.55 |
| Ah=A | 320 | 0-15 | 3.6 | 2.36 |
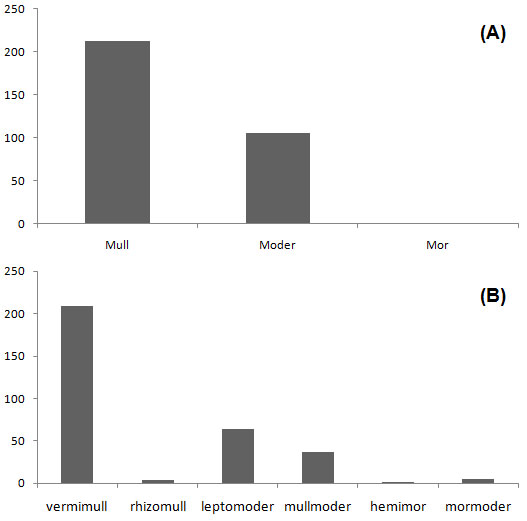
Using the key by Green et al. ([8]) Mull was the dominant humus form unit, with 213 of all 320 studied profiles (66.5%), Moder followed with 106 profiles (33.12%) and only 1 profile belonged to Mor (Fig. 1a). Six humus form subunits were observed and among them vermimull was the most widespread (209 samples, 65.3%) was the most dominant humus suborder while hemimor was the rarest one (1 sample). Other subunits found in the study site were leptomoder 64 (20%), mullmoder 37 (11.56%), mormoder 5 (1.56%) and rhizomull 4 (1.25% - Fig. 1b).
Fig. 1 - Abundance of humus main references (a) and subunits (b) according to Green et al. ([8]).
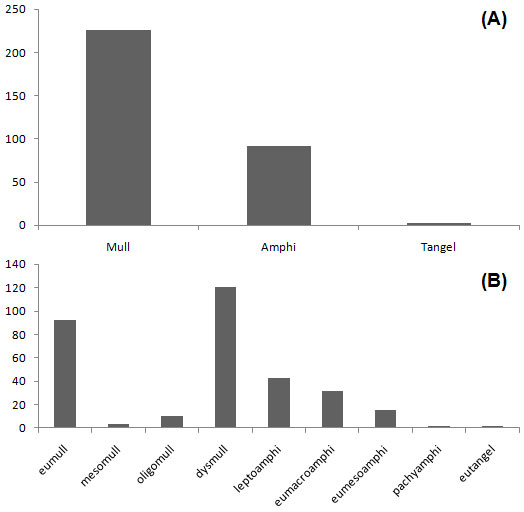
Using the classification of humus forms by Zanella et al. ([19]), Mull was the most widespread humus form unit, with a frequency of 226 (70.62% of all samples) in the study site. Amphi and Tangel with the frequencies of 92 (28.75%) and 2 (0.62%), respectively. Nine subunits were found in the study site: dysmull, with 121 samples (37.1%) and eumull, with 92 samples (28.75%), were the commonest humus form subunits, while pachyamphi and eutangel with 2 samples each (0.62%) were the scarcest humus form subunits in the study site (Fig. 2).
Fig. 2 - Abundance of humus main references (a) and subunits (b) according to Zanella et al. ([19]).
Discussion
The abundance of Mull in the study site, 66% using Green et al. ([8]) and 71% using Zanella et al. ([19]), indicates a decomposition rate and a rapid return of foliage nutrients into the soil ([2], [20]). It also shows good habitat conditions for soil organisms such as suitable aeration, balanced moisture supply, nutrient-richness, proper temperature and an input of easily decomposable litter ([8]).
A previous study by Sajedi et al. ([15]) revealed that Moder was the dominant humus form in some pure beech stands of Hyrcanian forests while the present study, realized in a mixed broadleaved part of Hyrcanian forests, shows the dominance of Mull humus forms. This difference is probably due to the difference in species composition of the canopy. The presence of Mull-forming tree species like hornbeam (Carpinus betulus) within a stand dominated by Moder-forming species such as beech (Fagus orientalis) has probably a positive impact on the forest floor decomposition rate and can facilitate the building of a Mull form. A similar impact of mixed canopies has already been observed by Brandtberg et al. ([5]) and Aubert et al. ([1]).
According to Green et al. ([8]), Mull is a humus form in which organic matter is combined with the upper mineral soil instead of being accumulated on its surface as in Mor and Moder humus forms. Actually, the most common Mull subunit in the study site is vermimull, resulting from a high activity of large earthworms, which are able to incorporate organic matter and mineral particles, forming the topsoil crumby structure of Mull. This also proves that trees in the studied stands produce easily decomposable litter. The next most abundant humus subunits were leptomoder and mullmoder, both belonging to the Moder unit according to these authors. These two subunits encompass a rapid decomposition rate and a high soil faunal activity, but this activity is not high enough to completely decompose and make disappear organic horizons as this occurs in the Mull unit ([8]).
According to Zanella et al. ([19]), Mull was the dominant humus form unit in the study site. Mull is an indicator of a temperate climate condition with high soil fauna activity and rapid decomposition rate ([10], [15], [20]). This humus form occurs in stands that produce easily biodegradable litter with relative low C/N ratio and without serious environmental constraints. The pHwater of the A horizon in Mull order is often more than 5 ([20]), in agreement with a pH ranging from 5.3 to 7.8 performed by Habashi ([9]) in the same site.
Concerning terrestrial ecosystems (never or few days per year submerged or water saturated soils), the employed keys of classification are able to identify 3 (Canadian: Mull, Moder and Mor) or 5 (European: Mull, Moder, Mor, Amphi and Tangel) main references. European Amphi, Tangel, Moder and Mor correspond to Canadian Moder or Mor. In fact, considering the influence of the substrate, the European classification shares calcareous and siliceous topsoils. On calcareous substrates Amphi corresponds to a twin humus form showing both characters of Mull (crumby organo-mineral horizon) and Moder (accumulated organic humus), due to a seasonal alternation between phases of high and low biological activity in strongly seasonal Alpine and Mediterranean environments ([11]). Tangel expresses particular characters at high elevation and on hard calcareous rocks, where litter is out of reach of soil decomposer activity for most of the year and invertebrates cannot dig through the parent rock. In both European and Canadian classifications, starting from a neutral Mull, Moder and Mor correspond to a scale of decreasing nutrient availability and colder conditions, stemming in decreasing biological diversity and activity on acid substrates.
The main references are parted in a second level of 16 subunits in Canadian or European classification as well. In the study area, 6 or 9 subunits were detected with Canadian ([8]) or European ([19]) keys respectively. The Green et al. ([8]) reference comprises only 3 Mull subunits while the Moder and Mor enumerate 6 and 7 subunits respectively. This means that the Canadian key emphasizes Mor and Moder humus forms which are common in this country. On the other hand the European key encompasses of 4 Mull, 4 Amphi, 3 Moder, 2 Tangel and 3 Mor subunits. Regarding the temperate climate condition of the investigated site, Mull, Amphi and Moder (or Mull and Moder in [8]) are more expectable than Mor forms. Leptoamphi sub-unit in European method mostly correspond to vermimull and leptomoder sub-units in Canadian method, eumacroamphi correspond to leptomoder, vermimull and mullmoder, eumesoamphi correspond to leptomoder, pachyamphi correspond to hemimor, eutangel correspond to mormoder sub-unit in as well. This correspondence is mainly due to similar definition of these four units (Amphi, Moder, Tangel, Mor):
- Amphi or Moder: According to the European classification Amphi order encompasses humus profiles in which there is "simultaneous presence of OH, A biomacro or A biomeso horizons, absence of OFnoz, thickness of A horizon ≥ thickness of ½ OH horizon, absence of A massive or single grain, presence of A biomacro and one of the following; living earthworms in the A horizon, sharp transition between A and OH, pH in water of the A horizon ≥ 5". While in Moder according to the aforementioned classification following characteristics are expected: "presence of OH horizon, absence of OFnoz, absence of A biomacro, absence of A biomeso and one of the following, no sharp transition OH/A horizon (transition ≥ 5 mm), pH in water of the A horizon < 5, presence of A biomicro". But Canadian classification does not mention neither an especial pH nor an A to OH transition condition for Moder order, and emphasize on the horizon thickness and zoogenous or fungal activities as the most important criteria to classify the humus profiles.
- Tangel or Mor: According to the European classification a Tangel order must has the following characteristics: "presence of thick organic zoogenic horizons (OFzo + OH > 5 cm), hard limestone and/or dolomite rock/rock fragments at the bottom of the humus profile, cold climate (subalpine or upper montane belts), absence of OFnoz, presence of a thin (thickness < ½ OH) A massive or single grain or biomeso; or pH in water of the a thin (thickness < ½ OH) A horizon ≥ 5". In this study site only two Tangel orders were found in some rare stone outstanding (mostly sand stone) scattered out in the area. But Mor order is characterized by a mass accumulation of organic matter and a thick Fm (mycogenous F horizon) and zoogenous activity is very low ([8]). In the study site only one humus profile hardly were found having (almost) such characteristic which is negligible.
Considering all above facts it can be concluded that in comparison between two main humus classification methods, the European key classification reveals itself more precise and seems more suitable than the Canadian one for mixed oriental beech forests and similar ecosystems. The same category (such as Mull or Moder) did not gather an equal number of soil profiles with European or Canadian classification. This is due to the difference in definition of units and sub-units in the classifications. A soil profile may be considered as a Mull (dysmull) in European method while in as Moder (mullmoder) humus form unit because two mentioned classifications emphasize different criteria in the definition of the main references and subunits. European method emphasizes the structure of A, pH and H to A transition condition as critical criteria while Canadian method mostly stresses the horizons thickness more that other characteristics of humus profile.
As a conclusion, the European humus classification method ([19]) is recommended for Hyrcanian mixed beech forests and similar ecosystems.
References
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
H Habashi
Faculty of Forest Sciences, Gorgan University of Agricultural Sciences and Natural Resources, Gorgan (Iran)
Corresponding author
Paper Info
Citation
Waez-Mousavi SM, Habashi H (2012). Evaluating humus forms variation in an unmanaged mixed beech forest using two different classification methods. iForest 5: 272-275. - doi: 10.3832/ifor0632-005
Academic Editor
Alberto Santini
Paper history
Received: Dec 04, 2011
Accepted: Sep 27, 2012
First online: Oct 24, 2012
Publication Date: Oct 30, 2012
Publication Time: 0.90 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2012
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 58654
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 49217
Abstract Page Views: 3256
PDF Downloads: 4552
Citation/Reference Downloads: 15
XML Downloads: 1614
Web Metrics
Days since publication: 5008
Overall contacts: 58654
Avg. contacts per week: 81.98
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2012): 8
Average cites per year: 0.57
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Dynamics of humus forms and soil characteristics along a forest altitudinal gradient in Hyrcanian forest
vol. 14, pp. 26-33 (online: 10 January 2021)
Research Articles
Influences of forest gaps on soil physico-chemical and biological properties in an oriental beech (Fagus orientalis L.) stand of Hyrcanian forest, north of Iran
vol. 13, pp. 124-129 (online: 07 April 2020)
Research Articles
Potential relationships of selected abiotic variables, chemical elements and stand characteristics with soil organic carbon in spruce and beech stands
vol. 14, pp. 320-328 (online: 09 July 2021)
Research Articles
Classification and mapping of Spanish Mediterranean mixed forests
vol. 12, pp. 480-487 (online: 14 October 2019)
Research Articles
Dynamics of soil organic carbon (SOC) content in stands of Norway spruce (Picea abies) in central Europe
vol. 11, pp. 734-742 (online: 06 November 2018)
Research Articles
Fine root morphological traits and production in coniferous- and deciduous-tree forests with drained and naturally wet nutrient-rich organic soils in hemiboreal Latvia
vol. 16, pp. 165-173 (online: 08 June 2023)
Research Articles
The effect of clear-cut age on soil organic carbon and nitrogen indices in Scots pine (Pinus sylvestris L.) stands
vol. 18, pp. 146-153 (online: 09 June 2025)
Research Articles
Short-term effects in canopy gap area on the recovery of compacted soil caused by forest harvesting in old-growth Oriental beech (Fagus orientalis Lipsky) stands
vol. 14, pp. 370-377 (online: 10 August 2021)
Review Papers
The soil-conscious forestry and the forbidden apple
vol. 17, pp. 252-268 (online: 16 August 2024)
Short Communications
Not all long-distance-exploration types of ectomycorrhizae are the same: differential accumulation of nitrogen and carbon in Scleroderma and Xerocomus in response to variations in soil fertility
vol. 14, pp. 48-52 (online: 18 January 2021)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







