
Plant cover evolution and naturalisation of revegetated ski runs in an Apennine ski resort (Italy)
iForest - Biogeosciences and Forestry, Volume 2, Issue 5, Pages 178-182 (2009)
doi: https://doi.org/10.3832/ifor0511-002
Published: Oct 15, 2009 - Copyright © 2009 SISEF
Research Articles
Abstract
Revegetation on ski runs is performed to limit soil erosion, to reduce visual impact and to lengthen the duration of snow cover. Commercial forage mixtures are often used and the mid-long term aim is to obtain a high level of colonisation by species of the local flora to ensure the ecological integration of restored areas. The naturalisation of the artificial canopy is affected in a remarkable way by the local environmental conditions and by management of the swards. To contribute to the knowledge of this process, data collected in an Apennine ski resort on several ski runs revegetated in different years and located at different elevations (from 1020 to 1745 m a.s.l.) are reported. Botanical analyses were also conducted on natural areas located at different altitudes and close to the ski tracks to assess the level of naturalisation reached by the artificial canopy. Results showed the effectiveness of the revegetations performed, even though they were carried out with very simple mixtures. Data collection allowed to evaluate the rapidity of the naturalisation process which is mainly affected by the height of the ski slope and years since sowing. Evolution of swards over time demonstrated that, in the environment studied, characterised by low altitude, the speed of colonisation by native species is considerably higher than that reported in previous studies carried out in different alpine environments located at upper elevations.
Keywords
Revegetation, Botanical composition, Native species, Floristic richness
Introduction
Winter tourism today represents one of the most important economic sectors in a great part of the world’s mountain areas ([9], [25]). At the same time, the impact of ski runs and other infrastructures related to ski sports may have dramatic effects on fragile mountainous environments ([23], [29], [22]). As a consequence, many studies have been carried out on the restoration of ski slopes after their creation ([31], [29], [25]), as their construction is one of the major causes of anthropogenic disturbances in alpine ecosystems ([35], [16]). If the analysis is limited to the ski run routes, the major negative effects are on soil and vegetation, which are often affected by any human disturbance to ecosystems ([33], [11], [15]). Soils are disturbed by the construction of ski runs and ski-lifts, by skiers’ passage and snowcats managing the snow ([10]). Damage caused by skiers is more clearly seen in the middle of the ski runs ([24]), where the machines create undesirable effects over the entire soil surface. The high level of snow compaction produces a denser snow layer, with a reduction in snow insulating properties, a higher probability of soil freezing, decreasing gas permeability, alteration of bio-geochemical cycles, and a decrease in micro-porosity (e.g. [12], [36]). All of these changes can dramatically affect the vegetation of ski runs ([25]).
Taking into account these considerations and the environmental constraints in fragile alpine environments, revegetation is always performed after the construction of a ski run in order to limit soil erosion, reduce visual impacts, and maintain and lengthen the duration of snow cover ([2]). Rapid reconstitution of vegetation cover is desirable in order to reach these goals but many studies carried out in Alpine Mountain areas (e.g., [30], [8], [1], [27]) have shown the difficulty and slowness of this process, especially in areas higher than 1800-2000 m a.s.l., such as those above the timberline ([6], [32]). Commercial mixtures composed of forage species are generally used for revegetation because they are cheap and easily available ([3], [14]), but these species are often not suitable to local conditions ([26], [32], [13]). Therefore, the use of seed from local natural populations ([19]) or from species adapted to the intervention site is recommended ([21]) to reduce failures and to speed up the establishment of a stable plant community. For this reason it is advisable to conduct programs for the preliminary evaluation of the species to be used in several areas so as to create a suitable mixture of species (sometimes not conventionally used mixes) well adapted to particular pedo-climatic conditions ([2]). Once the revegetation has successfully been performed, the mid-long term aim is to obtain a high level of colonisation by native species to assure the ecological integration of restored areas ([8], [1]). Bédécarrats ([6]) identified three stages of development of these plant communities: a beginning stage, in which sown species dominate the sward, a transitional stage, in which the colonisation of the ski run by native species begins to replace the sown plants, and a maturation stage, in which the proportion of autochthonous species is higher than those introduced by revegetation. Talamucci & Argenti ([28]), analysing data from 21 ski slopes scattered across 12 Italian resorts, found similar patterns of recovery by native species and stated that on average 6 to 9 years are needed to reach a significant amount of autochthonous plant presence (30-40% out of the total) in the study areas. The rate of recolonisation by autochthonous species is affected by management and local environmental conditions.
To contribute to the knowledge of vegetation development in revegetated ski runs and to clarify the rate of recovery by native species, a research was carried out in a Northern Apennine ski area using data from several ski runs that had been restored in different years.
Materials and methods
Research was carried out at the Monte Cimone ski resort (North Apennines, Italy, 44° 11’ N, 10° 42’ E), a ski area with 30 ski runs totalling about 50 km of tracks. In this area eight different ski slopes ranging in elevation between 1020 and 1745 m a.s.l. were identified (Tab. 1). These ski slopes were revegetated in different years (from 1993 to 2004) and it was possible to recover the constituents of the seed mixes used. Ski slopes were revegetated by hydroseeding utilising a seeding rate of 400-500 kg ha-1 and simple grass/legumes mixtures (2-3 species). Festuca gr. rubra L. was the most abundant component, contributing 80% of seeds by weight in all mixes. Legumes used were Trifolium repens L. (20% in three mixtures) and, in one case, Lotus corniculatus L. The vegetation was typically managed through one midsummer cutting. In the summers 2003-2004 a botanical survey was conducted on the established swards of the ski slopes whose age ranged from 1 to 11 years. Sixty botanical relevés were performed on the studied tracks, with a sampling intensity of about 100-150 m along each ski run. Data were collected by linear analysis according to the Daget & Poissonet ([7]) method on 25m transects with a quadrat every metre. The data collected made it possible to obtain the specific contribution (SC - the percentage of the presence of each species) and to assess the number of species for each transect in order to describe the existing floristic richness. Knowledge of the composition of the seed mixes allowed to distinguish between sown species and species deriving from natural colonisation. At the same time, the ground cover was observed by visual estimation for each transect. Due to their proximity, ski runs revegetated in 2000 were considered in the analysis as a single group called “Passo del Lupo”. The “Nord Beccadella” ski run was only examined in 2004 as its revegetation was carried out in spring of the same year.
Tab. 1 - Description of studied ski slopes: year of revegetation, mixture and seed rate used, elevation and number of transects performed.
| Ski slope | Year of revegetation |
Mixture used |
Seed rate (kg ha-1) |
Elevation (m a.s.l.) |
Number of plots |
|
|---|---|---|---|---|---|---|
| 2003 | 2004 | |||||
| Direttissima | 1993 | Festuca gr. rubra L. (80%), Trifolium repens L. (20%) | 400 | 1505-1730 | 7 | 4 |
| Rossa | 1995 | Festuca gr. rubra L. (80%), Trifolium repens L. (20%) | 400 | 1020-1350 | 6 | 4 |
| Colombaccio | - | - | - | - | 2 | 4 |
| Raccordo Cimoncino |
- | - | - | - | - | 3 |
| Lago della Ninfa |
2000 | Festuca gr. rubra L. (80%), Trifolium repens L. (10%), Lotus corniculatus L. (10%) | 500 | 1360-1660 | 5 | 3 |
| Betulle | - | - | - | - | 5 | 3 |
| Sud Beccadella | - | - | - | - | 6 | 4 |
| Nord Beccadella |
2004 | Festuca gr. rubra L. (80%), Trifolium repens L. (20%) | 400 | 1550-1745 | - | 4 |
In two areas located at 1700 m and 1300 m a.s.l., external but close to the ski tracks, five transects were performed using the same procedures described above in order to study the native vegetation in less disturbed areas and to compare the natural botanical composition to that deriving from ski run recovery. For this comparison, the data used for the ski tracks were those taken from the Direttissima and Rossa slopes, which were the oldest and the closest to these natural areas. Similarity of botanical composition among sown and natural sites was assessed by two indices based on species presence and on both species and abundance according to Warren et al. ([34]). The first is the Jaccard index ([18]), which measures the coefficient of similarity between two sample sets of data by the following formula (eqn. 1):
where c is the number of species common to both sites, and n and r the number of species found in natural and revegetated sites respectively. Thus, the comparison is carried out only on the floristic lists of the two areas, without taking into account the abundance of each species. The second index is the following, called the Similarity Index (SI - [5] - eqn. 2):
where |di| is the absolute value of the difference between the SC of the i-species in natural and revegetated areas. Following these formulas, the Jaccard and SI indices theoretically range from 0, when natural and revegetated areas are completely different (i.e., no common species), to 1, when the vegetation of the two areas are completely equal (both by species presence and their specific contributions).
Statistical analysis was performed by SYSTAT software using General Linear Model procedure to compare data from different slopes. Groups were compared using the Tukey test.
Results and Discussion
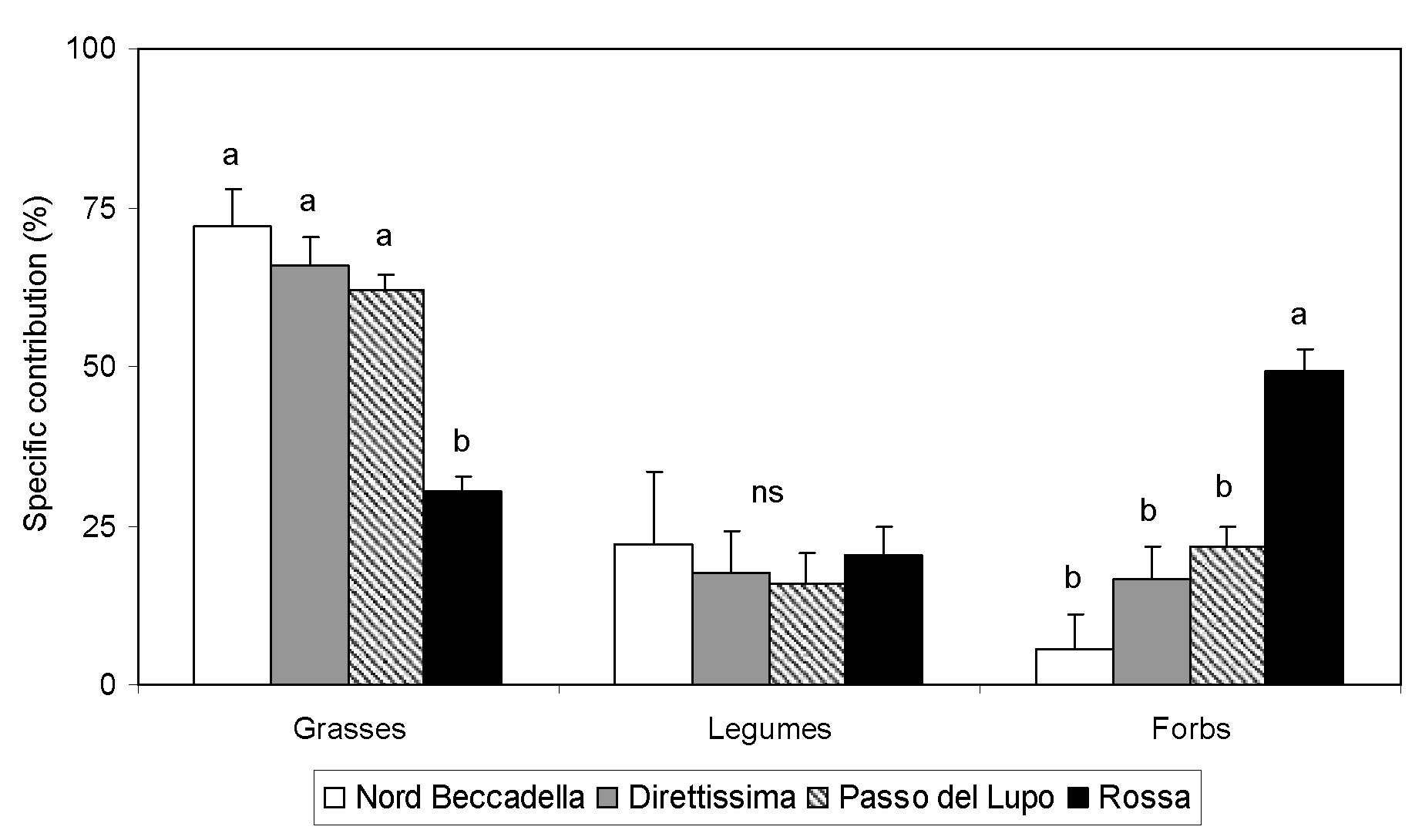
Specific contributions of species for all studied ski slopes were grouped by grasses, legumes and forbs and presented in Fig. 1 sorted from the highest elevation ski slope (Nord Beccadella) to the lowest elevation (Rossa). Grasses dominated almost all the slopes, except that located at the lowest elevation, which is also one of the oldest. Legumes occurred on all the slopes with similar and low values, and forbs were characterised by a reduced contribution, with the exception of Rossa where their SC was a bit less than 50% of the total.
Fig. 1 - Specific contribution for grasses, legumes and forbs in each ski slope. Values with the same letter are not significantly different (p<0.01, Tukey test, ns = not significant). Vertical bars represent standard errors.
Ground cover, botanical composition and number of species by sown and native species are reported in Tab. 2. Soil coverage was particularly low in Nord Beccadella as measures were taken in the same year of revegetation; the other ski runs attained remarkable values close or higher to the threshold of 70% of ground cover, which is considered the limit to be reached to reduce soil erosion ([17]). Sown grasses dominated the most recent revegetated ski run (Nord Beccadella) as legumes were poorly represented in almost all studied tracks, also taking into account the original composition of the mixtures used. Native species were dominant in the oldest ski slopes and native grasses were the prevalent group in the upper tracks (Direttissima, Passo del Lupo), and in the lowest (Rossa) naturalisation was mainly performed by forbs. As for the total number of species observed during the botanical survey, all the sown species were found even in the oldest tracks, as the native species were a relevant number in all the ski runs with the exception of Nord Beccadella, where only 3 autochthonous species occurred. In total, 99 different species were recorded along all the studied tracks.
Tab. 2 - Ground cover, botanical composition and number of species for sown and native species in the studied ski runs. Values in a column with the same letter are not significantly different (p<0.01, Tukey test).
| Ski slopes | Year of Revegetation |
Elevation (m a.s.l.) |
Ground cover (%) |
Sown species | Native species | No. of observed species | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Grasses | Legumes | Grasses | Legumes | Forbs | Sown | Native | Total | ||||
| Direttissima | 1993 | 1505-1730 | 77a | 22.0b | 10.3a | 43.9a | 7.3b | 16.5b | 2 | 49 | 51 |
| Rossa | 1995 | 1020-1350 | 86a | 14.4c | 2.1b | 15.9b | 18.4a | 49.2a | 2 | 55 | 57 |
| Passo del Lupo | 2000 | 1360-1660 | 69a | 31.8b | 10.6a | 30.5a | 5.3b | 21.8b | 3 | 53 | 56 |
| Nord Beccadella | 2004 | 1550-1745 | 25b | 66.2a | 5.8b | 5.4c | 16.6a | 6.0c | 2 | 3 | 5 |
Vegetation was thus strongly influenced in the younger sward by mixture species, particularly Festuca gr. rubra L., but this species was found on all ski slopes, demonstrating its persistence even after some years since sowing. The presence of forbs, all represented by native plants, indicated an ongoing colonisation process which was stronger in the lowest ski slope. Native species occurring on the slopes showed different behaviour. Some species, such as Geum montanum L., Dianthus deltoides L. and Euphrasia minima L., were more common at higher altitudes and on less disturbed tracks, and other species, such as Poa trivialis L., Anthriscus sylvestris (L.) Hoffm. and Galium mollugo L., occurred with high SC in lower tracks and in areas characterised by a higher anthropogenic disturbance.
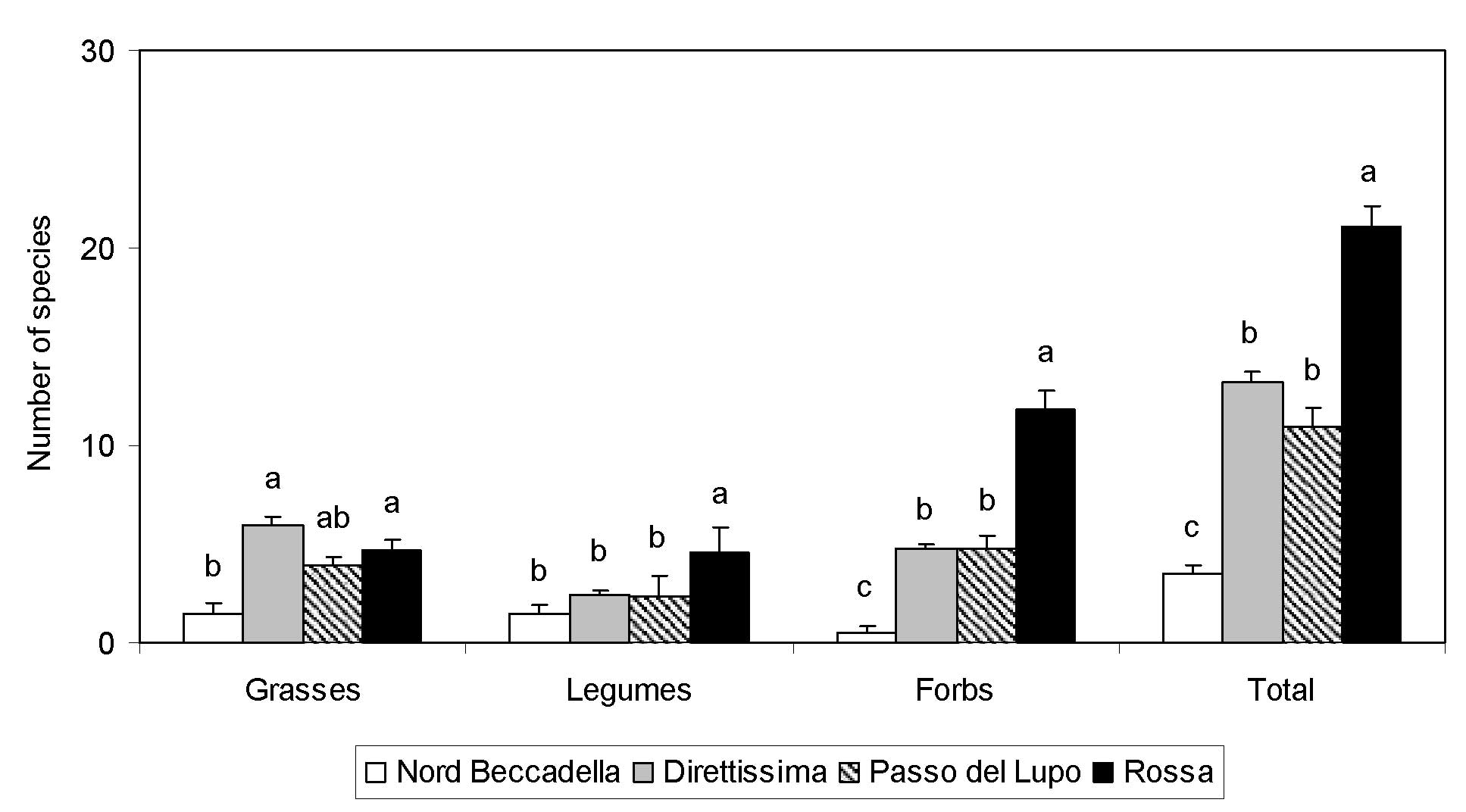
The mean number of species recorded along a linear analysis, grouped by grasses, legumes and forbs, and total, are shown in Fig. 2. Total numbers were very low in Nord Beccadella due to the very young age of the vegetation, and they were very high in Rossa, where the floristic enrichment of the sward was influenced by the low elevation and time since sowing. The number of grass species was positively influenced by vegetation age and in some situations grass richness was related to vegetation development. The presence of grasses such as Arrhenatherum elatius (L.) Presl, Festuca pratensis Huds. and Lolium perenne L. in the lower track testified that these species colonized the ski run from nearby grasslands used for forage production. The mean number of legumes and forbs was influenced by ski slope elevation since their presence increased significantly in lower elevation areas. The number of forbs collected along the tracks is the main variable affecting overall floristic richness.
Fig. 2 - Average number of species along a transect for grasses, legumes, forbs and their totals in each ski slope. Values with the same letter are not significantly different (p<0.01, Tukey test). Vertical bars represent standard errors.
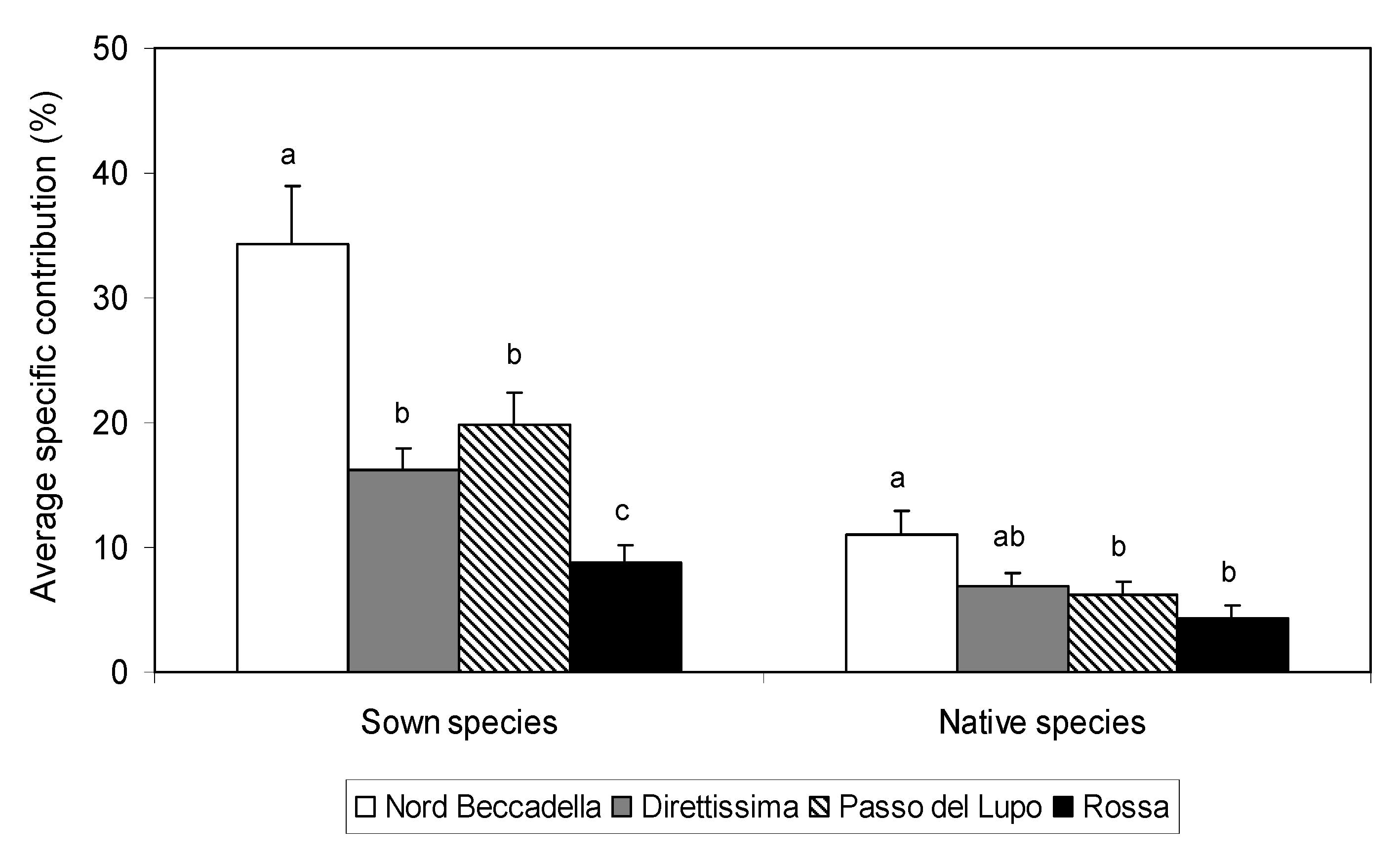
To evaluate the overall presence of particular species categories, we calculated the average specific contribution separately for sown and native species (Fig. 3). Occurrence of sown species is mainly influenced by number which is almost the same in all ski slopes due to their persistence after sowing. The highest value of average specific contribution for sown species was found in the youngest swards (i.e., Nord Beccadella), that were dominated by a low number of species introduced with sowing. The lowest value is that of Rossa ski slope in which the colonization process reduced the overall specific contribution of sown species recorded on the track. The mean contribution by native species was always lower than sown species and this was strongly influenced by ski slope elevation. In the lower areas native species occurring on the tracks were present with a high number of species but each of them was represented by a very low specific contribution. By comparing the study sites it is possible to postulate that the difference between the average specific contribution of sown and native species is a useful parameter for estimating the level of development of naturalisation for artificial vegetation.
Fig. 3 - Average specific contribution for sown and native species in each ski slope. Values with the same letter are not significantly different (p<0.01, Tukey test). Vertical bars represent standard errors.
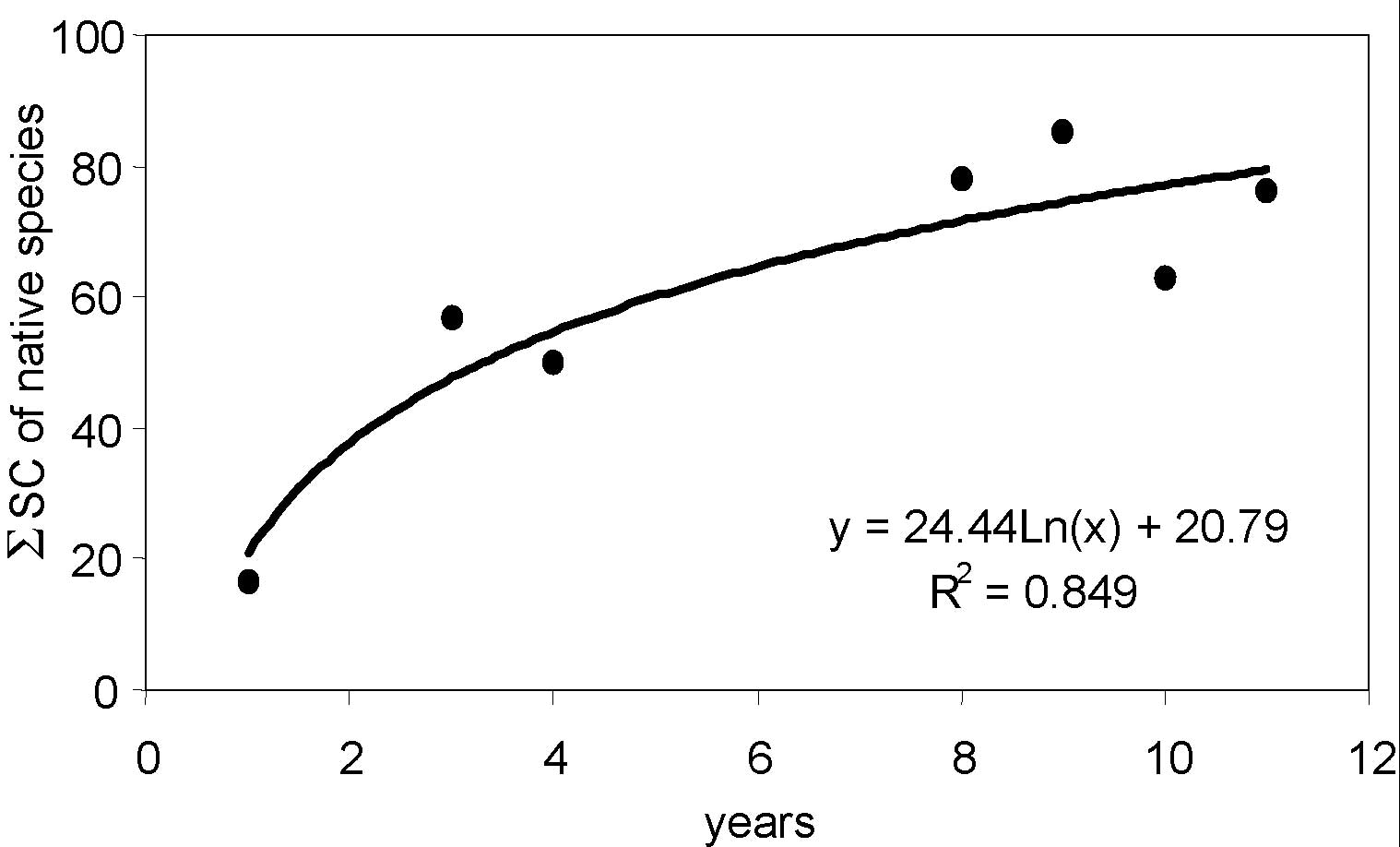
Native species recovery as a function of years since sowing is described in Fig. 4. The regression model was a logarithmic one, so we can state that the speed of recolonisation, measured in terms of SC of native species, was higher in the early years but later slowed as described for alpine environments by Argenti et al. ([1]) and Talamucci & Argenti ([28]). It may not be possible to obtain a complete naturalisation of ski slopes due to the long persistence of sown species.
Fig. 4 - Relationship between overall specific contributions of native species and number of years since sowing.
Taking into account that ski run elevation and time since revegetation are the main factors affecting the naturalisation process, both the above parameters were used as predictors in multiple regression to explain the presence of native species. The resulting equation is the following (eqn. 3):
where SC ns is the total amount of specific contributions of all native species, h is the elevation of the ski run (m a.s.l.) and a is the number of years since revegetation. Therefore, for each year following revegetation, the SC of native species increases by about 4% and for each 100 m gain in altitude, the SC of native species decreases by about 4%.
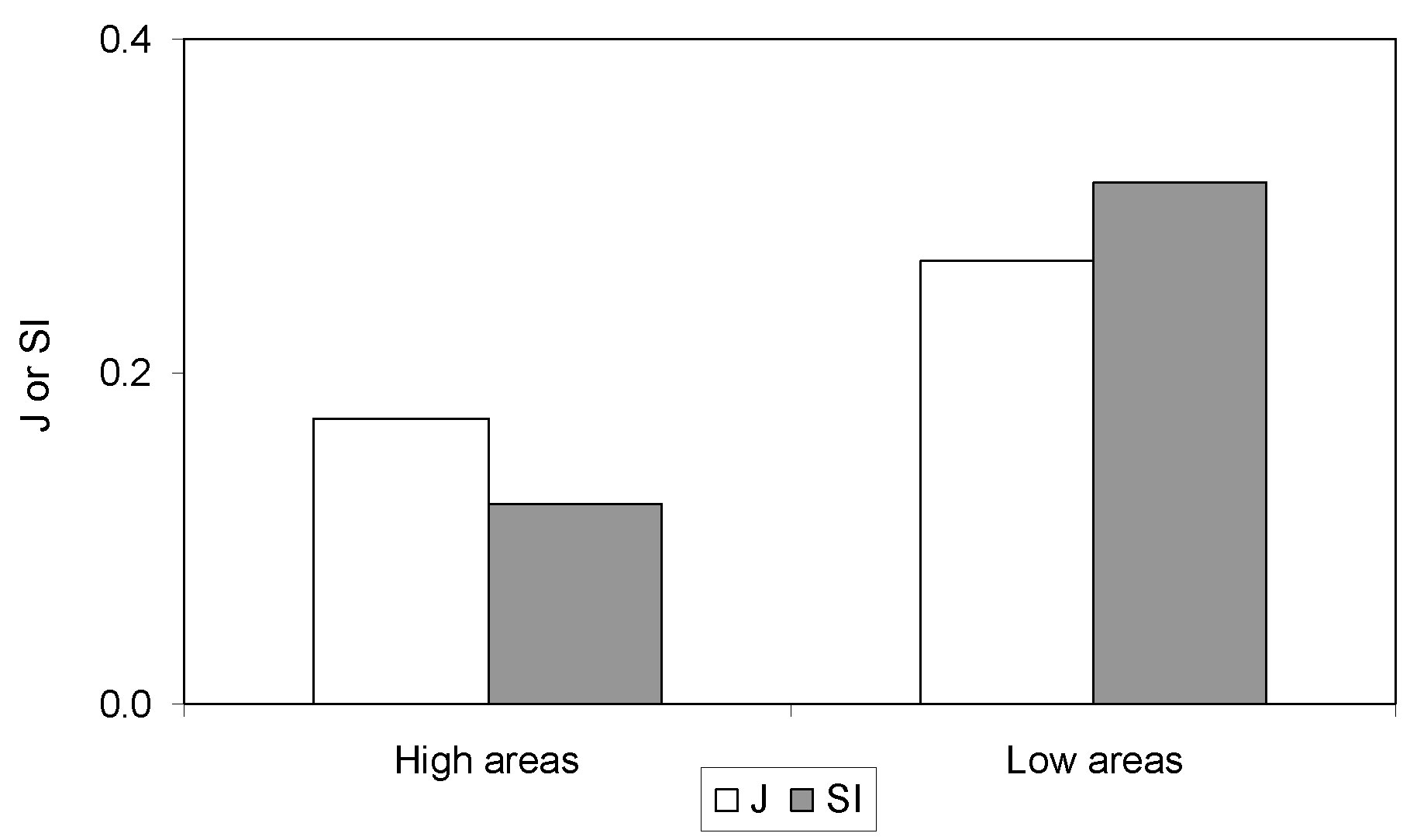
A comparison of species composition in revegetated ski slopes and nearby natural areas located at different heights was carried out using the described similarity indices (Fig. 5). The number of years since sowing and seed mixtures used for the ski slopes involved in this particular comparison were similar, so differences in vegetation composition for this case were ascribed to elevational effects. Even if index values for the same area were slightly different for J and SI, this would be due to the different algorithm of calculation and they both discriminated high elevation (about 1700 m a.s.l.) from low elevation (about 1300 m a.s.l.) areas. Higher values in the lower elevation sites showed that altitude was very important in the level of naturalisation achieved by the vegetation, both in terms of number of species and their contribution.
Fig. 5 - Average Jaccard index (J) and Similarity Index (SI) values for the high and low elevation areas.
Conclusions
Revegetation of ski slopes can be carried out utilising different technical choices with the mid-long term objective of the reconstitution of stable and semi-natural vegetation. A major role in such naturalisation can be played by the species belonging to autochthonous flora that recover the disturbed tracks through different mechanisms. This process is affected by some environmental conditions (air and soil temperature, duration of growing season, snow melt, air moisture, etc.), which could be synthetically summarised by the altitude. The timberline is usually considered as the limit above which the climatic conditions determine remarkable difficulties for restoration ([21]). In any case, it is hard to achieve good results in restoration at high altitude relying only on recovery of native species ([20]). Therefore, it is advisable to revegetate these disturbed environments and then control the naturalisation process.
In our study the recovery of native species was favoured by the low average altitude of the studied ski runs. For this reason, the speed of naturalisation was higher than that found in areas located at upper elevation (e.g., [6], [28]). Actually, our data showed that on average about 4 years were necessary after sowing to reach an equal proportion between sown and native species. According to Bédécarrats ([6]), this may be considered the threshold of the achivement of a maturation stage along the evolution process. These findings demonstrate that the constitution of semi-natural plant communities can be achieved in a short period also by sowing commercial mixtures without the use of particular autochthonous species.
Results showed that site altitude and time since sowing are the most important variables that can be used to describe the recovery performed by native species. In addition, a simple multiple regression model considering only these two parameters can explain a high proportion of the variability of the presence of native species, confirming what found by Argenti et al. ([4]) for ski slopes in alpine environments.
Moreover, the analysis of average specific contribution made it possible to attribute a separate role to different categories of species. Thus, while sown species are considerable in terms of specific contribution, they are few in number, and can carry out mainly a defensive and protective function. On the other hand, native species, which are significant in number but low as proportion, can play mainly an ecological role and contribute to the floristic enhancement of the artificial swards in a better way.
Acknowledgements
Authors contributed in equal parts. Authors thank the staff of the Monte Cimone ski resort and Dr. G. Betta for their support. Research supported by Italian Ministry of Agricultural and Forestry Policies.
References
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Corresponding author
Paper Info
Citation
Argenti G, Ferrari L (2009). Plant cover evolution and naturalisation of revegetated ski runs in an Apennine ski resort (Italy). iForest 2: 178-182. - doi: 10.3832/ifor0511-002
Academic Editor
Marco Borghetti
Paper history
Received: Mar 17, 2009
Accepted: Jun 26, 2009
First online: Oct 15, 2009
Publication Date: Oct 15, 2009
Publication Time: 3.70 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2009
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 48452
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 42472
Abstract Page Views: 2082
PDF Downloads: 3173
Citation/Reference Downloads: 21
XML Downloads: 704
Web Metrics
Days since publication: 5308
Overall contacts: 48452
Avg. contacts per week: 63.90
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Feb 2023)
Total number of cites (since 2009): 4
Average cites per year: 0.27
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Scale dependency of the effects of landscape structure and stand age on species richness and aboveground biomass of tropical dry forests
vol. 16, pp. 234-242 (online: 23 August 2023)
Research Articles
Influences of mature Pinus nigra plantations on the floristic-vegetational composition along an altitudinal gradient in the central Apennines, Italy
vol. 13, pp. 279-285 (online: 03 July 2020)
Research Articles
Exploring patterns, drivers and structure of plant community composition in alien Robinia pseudoacacia secondary woodlands
vol. 11, pp. 586-593 (online: 25 September 2018)
Research Articles
Light availability influences the invasion of Teline monspessulana (L.) K. Koch in a temperate fragmented forest in Central Chile
vol. 15, pp. 411-416 (online: 19 October 2022)
Review Papers
Large-scale effects of forest management in Mediterranean landscapes of Europe
vol. 6, pp. 342-346 (online: 29 August 2013)
Research Articles
Agronomic methods for mountain grassland habitat restoration for faunistic purposes in a protected area of the northern Apennines (Italy)
vol. 9, pp. 490-496 (online: 07 January 2016)
Research Articles
Long-term outcome of precommercial thinning on floristic diversity in north western New Brunswick, Canada
vol. 1, pp. 145-156 (online: 25 November 2008)
Research Articles
Regeneration dynamics in the laurel forest: changes in species richness and composition
vol. 11, pp. 308-314 (online: 13 April 2018)
Research Articles
Effects of planted European beech on the understory in Scots pine forests of Lithuania
vol. 7, pp. 12-18 (online: 07 October 2013)
Research Articles
Shifts in the arbuscular mycorrhizal fungal community composition of Betula alnoides along young, middle-aged plantation and adjacent natural forest
vol. 13, pp. 447-455 (online: 07 October 2020)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







