Changes in organic compounds during leaf litter leaching: laboratory experiment on eight plant species of the Sudano-guinea Savannas of Ngaoundere, Cameroon
iForest - Biogeosciences and Forestry, Volume 1, Issue 1, Pages 27-33 (2008)
doi: https://doi.org/10.3832/ifor0450-0010027
Published: Feb 28, 2008 - Copyright © 2008 SISEF
Research Articles
Abstract
A laboratory experiment was carried out on the leaf litter of 8 agroforestry plant species of the Sudano-guinea Savannas of Ngaoundere in order to compare patterns of their water absorption and dynamics of four important energetic organic compounds (soluble sugars, cellulose, phenol and lignin) among these plant species during the leaching phase and to determine the influence of initial litter properties on these processes. To this end, 168 samples of leaf litter (5.00 ± 0.01 g) of Annona senegalensis, Lophira lanceolata, Syzygium guineense var. guineense, Syzygium guineense var. macrocarpum, Vitellaria paradoxa, Vitex doniana, Vitex madiensis and Ximenia americana were immersed for 15 days in distilled water at a temperature of 23 °C and relative humidity of 65 ± 1%. Three samples of each plant species were taken at 1, 6, 24, 72, 168, 240 and 360 hours. Depending on the species, water absorption capacity after 360 h of leaching varied from 162.77 (S. g. var. macrocarpum) to 264.00% (V. madiensis) of dry litter mass. The release of water-soluble substances varied between 9.61 (L. lanceolata) and 34.12% (X. americana). Water absorption and release of water-soluble substances rate constants were the highest in V. madiensis (0.32 h-1 and 0.25 h-1) and the lowest one respectively in S. g. guineense (0.03 h-1) and S. g. var. macrocarpum (0.006 h-1). Organic compound of original litter also varied significantly among species and decreased with leaching time for water-soluble sugars and phenols, while increased for cellulose and lignin. Water absorption by litter was significantly correlated with initial water-soluble sugars and phenol content, leaf litter area and thickness. The release of water-soluble substances by litters was also correlated with initial water-soluble sugars, water content and leaf litter area. The leaching rate constant was correlated with that of water absorption. These preliminary results lead to a better understanding of the litter decomposition processes of agroforestry plant species of Ngaoundere and thus help in making the most appropriate choice of indigenous plant species for domestication that could help to improve soil fertility and eventually to protect the biodiversity.
Keywords
Litter, Leaching, Quality, Organic compounds, Sudano-guinea savanna, Cameroon
Introduction
Litter decomposition represents an important process in the biogeochemical cycling. During this process immobilized nutrients in litter are released from litter to soil and made available to plants and soil micro-organisms ([34]). The litter decay rate is a factor that largely determines soil fertility ([8]) and its regulation plays an important role in agro-ecosystem functioning, notably in poor soil of Sudano-guinea savannas of Ngaoundere.
The initial phase of litter decomposition, so called leaching stage, has an important influence on subsequent processes and it may be responsible for a substantial part of mass loss from litter during the early stages of litter decomposition ([19], [27]). Bernhard-Reversat ([4]) reported that mass loss from fresh litter was equal to the loss of soluble compounds during the first weeks of in situ decay. This emphasized the dependence of the early decomposition processes on soluble organic compound release. Removal of soluble matter through leaching could thus impede or delay litter decomposition compared with litter in which soluble remain long enough to be metabolized. During the leaching phase, litter releases both inorganic elements, such as Ca, K, Mg, and simple organic compounds, such as protein and water-soluble sugars and phenols ([20], [6]). These are energetic compounds, necessary for the decomposers which will later decompose recalcitrant compounds such as lignin and cellulose ([3]).
Water-soluble compounds released by litter varies among species or specie groups ([14], [5]). It is also known that not all water-soluble compounds are leached with equal facility. Among the organic compounds, carbohydrates are easily and rapidly leached. They are generally easily and rapidly degraded in early stages of litter decomposition by fast growing microorganisms that may require a high concentration of nitrogen ([29]). Cellulose and lignin, the most abundant components of litter, are slowly decomposed. Nevertheless, because lignin physically protects most of the cellulose and hemicellulose from enzymatic hydrolysis, neither group of compounds decompose independently ([10]). High concentrations of polyphenols and lignin lead to low decomposition rate ([29], [18]).
However, in spite of the importance of the leaching phase in litter decomposition processes, information concerning leaching of organic compounds of litter of indigenous plant species of the Sudano-guinea savannas of Ngaoundere, particularly potential agroforestry plant species which have likely, in the future, to be integrated in farming systems in order to improve or restore soil fertility, is very limited, excepting that of Ibrahima et al. ([15]). Their study was carried out on water absorption and mass loss of nine litters during 240 h of leaching, and the relations between physical characteristics of leaf and leaching rates. The aim of the present study is to determine the dynamics of four important energetic organic compounds (soluble sugars, cellulose, phenol and lignin) during 360 h of litter leaching in laboratory, to compare the patterns of this process among these plant species and to determine relations between physical characteristics of litter and leaching.
Materials and methods
Leaf litter selection
In this study, only fresh fallen leaf litter of eight socio-economical plant species of the Sudano-guinea savannas of Ngaoundere was used. The experiment involved 8 species: deciduous broad-leaved including three tree species (Lophira lanceolata, Vitex doniana and Vitellaria paradoxa) and four shrub species (Annona senegalensis, Syzygium guineense var. macrocarpum, Vitex madiensis and Ximenia americana) and an evergreen broad-leaved tree species (Syzygium guineense var. guineense). The distribution area of S.g. var. guineense Engl. (Myrtaceae) and V. doniana Sweet (Verbenaceae) is the forest gallery, while A. senegalensis Pers. (Annonaceae), L. lanceolata Vann Tiegh ex Keay (Ochnaceae), S. guineense var. macrocarpum Engl. (Myrtaceae), V. madiensis Oliv. (Verbenaceae), V. paradoxa Vaertu (Sapotaceae) and X. americana Linn (Olacaceae) are located in the upland savanna. The eight species play a great socio-economical role. They are a source of income, food, firewood, medicinal substances and soil fertility indicators for the farmers of this region ([35], [25]). The farmers start now to conserve these plant species in farms.
Leaching experiment
New litter fall samples were collected directly from forest floor in the Ngaoundere humid savannas, next to the University of Ngaoundere, during maximum leaf fall period (November 2002 - January 2003). This period corresponds to dry season and soil was very dried; no leaching was occurred from new litter. The site, Dang, is located 15 km from Ngaoundere Town. Litter was air-dried and stored in the laboratory before use. For each species, twenty one leaf-litter samples of 5.00 ± 0.01g were weighed and submerged in plastic glass bottles (15 cm high and 13 cm wide), each containing 1 litre of distilled water, which was renewed every 24 h. The bottles were maintained in the laboratory for 15 days at room temperature of 23 °C and relative humidity of 65 ± 1%. Three other samples of each species were oven dried (48 h at 60 °C) to determine their initial water content and initial dry mass. Three sample replicates of each species were sampled from bottles after 1 h, 6 h, 24 h, 72 h, 168 h, 240 h and 360 h. After removal, the samples of each litter were weighed wet, after removing surface water by pressing them between two sheets of filter paper. Then they were dried (48 h at 60 °C) and re-weighed to obtain their dry weight. The water content and remaining dry mass were calculated.
For each species, 3 replicates (each of 10 leaf litter) were oven-dried (48 h at 60 °C), then weighed and their thickness measured by calliper. To avoid fragmentation, leaf-litter was moistened again, spread out and then the leaf areas were measured using a planimeter (Area meter, MK2).
Chemical analysis
The samples of dry initial litters and those collected after 168 h and 360 h of incubation (n = 72) were ground in Waring Commercial Blender and sieved at 1 mm. The concentrations of water-soluble sugar, phenol, cellulose and lignin were respectively determined by Dubois et al. ([11]) method, by Folin-Ciocalteu reagent ([21]), by colorimetric method ([32]) and by [33] and detergent method.
Statistical analysis
Changes in litter water content (WC in % dry mass) in relation to leaching time (in hours) were fitted to the following simple exponential model (e.g., [14] - eqn. 1):
where A is the maximum water absorption capacity of litter (or Asymptote), ke and t are respectively the litter water absorption rate constant and the time of water absorption.
The litter mass remaining (LMR expressed as a percentage of the initial mass) in relation to leaching time (in hours) was fitted to the following simple exponential model ([15], modified - eqn. 2):
where ks is the leaching rate constant, B and C are respectively the compartments of water soluble and resistant substances (lignin, cellulose and other compounds not determined).
Because their coefficients of determination were highly significant, both equations were adopted in study.
A multiple comparison among the fitted constants was carried out using the T’-method ([28]). ANOVA, following by Scheffe’s mean comparison test at 5% were used to compare the species.
The concentrations of the water-soluble sugars, phenol, cellulose and lignin compounds at the last sampling time (after 360 h of leaching) were compared to those measured in the initial litter for each species (Student t test). The loss of constituents was calculated as difference between final absolute amount and initial one of each component (g) and the same difference was also expressed as percentage of initial amount (%). A Student provided a test of the significance of the losses of constituents between the start of the experiment and after 360 h of leaching.
Pearson’s coefficients were calculated between water absorption and leaching rate constants and physical and chemical characteristics of initial leaf litter. Multiple regression (stepwise) models were also used to determine relationships between these parameters. Relationships between maximum water absorption and LMR after 360 h were also determined.
Results
Initial litter properties
Excepting lignin content, all initial litter properties presented in this study differed significantly among plant species (Tab. 1). Initial water content (IWC) varies from 4.12% in X. americana to 7.30% in S. g. macrocarpum. The highest values of leaf litter area (127.92 cm2) and thickness (0.47 mm) are found in A. senegalensis shrub and the lowest ones in X. americana shrub (15.92 cm2 and 0.14 mm). Water soluble sugar contents vary significantly from S. g. macrocarpum (5.90%) to V. paradoxa (12.63%) and those of phenol from V. madiensis (1.24%) to S. g. macrocarpum (3.69%). The lowest values of cellulose and lignin contents were found in litter of V. paradoxa (respectively 1.46 and 1.05%) and the highest ones were found respectively in those of V. doniana (3.28%) and L. lanceolata (5.37%).
Tab. 1 - Physical and chemical properties of initial litters (0h) and organic compounds of litter obtained after 360h of leaching. t: student test between litters at 0 h and 360 h of incubation; Standard Errors in parentheses; ns: non significant; ns: non significant; * P < 0.05; ** P < 0.01; *** P < 0.001. Different letters indicate significant differences among different species for initial litters or for litters after 360 h of incubation. AS: A. senegalensis; LL: L. lanceolata; SG: Syzygium guineense var. guineense; SM: S. guineense var. macrocarpum; VD: V. doniana; VM: V. madiensis; VP: V. paradoxa and XA: X. americana.
| Parameter | Time | Deciduous broad-leaved trees | Deciduous broad-leaved shrubs | Evergreen trees | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| LL | VD | VP | AS | SM | VM | XA | SG | F | ||
| Initial WC (%) | 0h | 5.27 (0.89)bc | 5.35 (0.15)c | 6.26 (0.84)bc | 5.14 (0.14)c | 7.30 (0.13)a | 4.90b (0.84)c | 4.12 (0.56)c | 6.46 (0.36)b | 6.58** |
| Area (cm2) | 0h | 116.08 (36.73)ab | 51.41 (36.46)c | 105.92 (23.24)ab | 127.92 (14.64)a | 95.83 (16.21)b | 43.36 (22.34)c | 15.92 (3.07)d | 84.67 (18.47)b | 21.10*** |
| Thickness (mm) | 0h | 0.21 (0.02)c | 0.40 (0.07)ab | 0.15 (0.03)d | 0.47 (0.04)a | 0.35 (0.03)b | 0.33 (0.05)b | 0.14 (0.03)d | 0.36 (0.04)b | 19.56*** |
| Soluble sugars (%) | 0 h | 7.91 (1.12)cd | 10.48 (0.03)ab | 12.63 (0.37)a | 9.54 (0.17)bc | 5.90 (1.14)d | 11.05 (0.97)ab | 10.99 (2.23)ab | 6.88 (0.84)d | 9.25** |
| 360h | 4.02 (0.11)bc | 4.15 (0.46)b | 2.46 (0.68)d | 5.86 (0.15)a | 3.81 (0.52)bc | 3.28 (0.20)bcd | 3.76 (0.14)bc | 3.01 (0.82)cd | 9.46** | |
| t | 4.91* | 19.22** | 18.67** | 22.92*** | 2.37ns | 11.13** | 4.58* | 4.66* | - | |
| Phenol (%) | 0 h | 1.84 (0.27)c | 1.54 (0.16)cd | 3.14 (0.34)b | 1.86 (0.14)c | 3.69 (0.36)a | 1.24 (0.09)d | 3.16 (0.09)ab | 3.21 (0.27)ab | 30.78*** |
| 360h | 0.99 (0.78)b | 0.55 (0.05)b | 2.17 (0.12)a | 0.73 (0.03)b | 0.84 (0.02)b | 0.16 (0.02)b | 2.13 (0.04)a | 0.71 (0.06)b | 6.92** | |
| t | 1.06ns | 8.58* | 3.75ns | 11.01** | 11.29** | 17.06*** | 14.86*** | 12.45** | - | |
| Cellulose (%) | 0 h | 2.53 (0.38)b | 3.28 (0.26)a | 1.46 (0.16)e | 2.02 (0.18)cd | 1.80 (0.17)cde | 2.16 (0.05)bc | 3.09 (0.09)a | 1.57 (0.14)de | 22.33*** |
| 360h | 3.35 (0.13)f | 6.55 (0.25)bc | 7.16 (0.09)a | 6.13 (0.02)d | 4.86 (0.10)e | 6.01 (0.17)d | 6.83 (0.17)b | 6.22 (0.10)cd | 146.85** | |
| t | 2.88ns | 12.80** | 44.22*** | 32.31** | 21.61** | 30.91** | 27.43** | 38.79*** | - | |
| Lignin (%) | 0 h | 5.37 (1.44)a | 3.25 (1.06)a | 1.05 (0.44)a | 3.47 (0.71)a | 3.22 (1.24)a | 2.27 (0.94)a | 3.01 (0.97)a | 2.57 (0.21)a | 3.29ns |
| 360h | 13.44 (0.88)a | 7.70 (0.33)cd | 4.69 (0.77)g | 9.76 (0.13)b | 6.15 (0.13)ef | 5.04 (0.97)fg | 6.94 (0.17)de | 8.37 (0.14)c | 51.88*** | |
| t | 6.77* | 5.69* | 5.78* | 12.33** | 3.34ns | 2.90ns | 5.66* | 32.59*** | - | |
Changes in water content of leaf litter
The WC of litter increased rapidly during the first 6 h of immersion, reaching 52.57 and 183.86% of dry mass respectively in S. g. guineense and V. madiensis (Fig. 1). Then the rate of increase slowed. At the end of incubation (15 days), the WC varied from 162.77% in S. g. macrocarpum to 264.51% in V. madiensis. Maximum water content (or maximum capacity of litter imbibition) was reaching after 72 h for the litter of A. senegalensis, L lanceolata, S. g. macrocarpum, V. paradoxa, V. madiensis and X. americana, because the values of water content among the four last sampling times were not significantly different (P > 0.05). Nevertheless, V. doniana and S. g. guineense reached their maximum water content respectively after 24 h and 240 h.
Fig. 1 - Changes in litter WC with leaching time in the 8 agroforestry species studied. AS: A. senegalensis; LL: L. lanceolata; SG: S.g. guineensis; SM: S. g. macrocarpum; VP: V. paradoxa; VD: V. doniana; VM: V. madiensis; XA: X. americana.
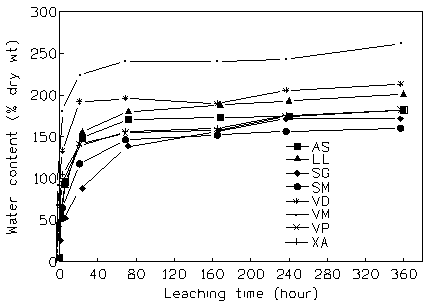
The water absorption was fitted to an exponential model for each litter type. All the fits were highly significant (Tab. 2). The absorption process seemed to have been completed at the end of 15 days, since the WC reached values which were not significantly different from the asymptote values (A) for all species (P < 0.05). The multiple comparison of asymptote values (A) and water absorption rate constants (ke) by T’-method a t 5% ([28]) showed significant differences among species (Tab. 2). V. madiensis was distinguished by the highest asymptote value and the highest absorption rate; both Syzygium species showed the lowest asymptote values, together with V. paradoxa and X. americana, and the lowest absorption rate constant.
Tab. 2 - Exponential regression equations describing changes in litter WC (in % dry mass) and LMR (in % of the initial mass) with leaching time (in hour). For each parameter, different letters indicate that the values are significantly different between plants species (P < 0.05, T’ method). All coefficients of determination (R2) are significant at P < 0.001. Standard Error in parenthesis and n = 21. AS: A. senegalensis; LL: L. lanceolata; SG: Syzygium guineense var. guineense; SM: S. guineense var. macrocarpum; VD: V. doniana; VM: V. madiensis; VP: V. paradoxa and XA: X. americana.
| Species | WC = A[1-exp(-ke t)] | LMR = B*exp(-ks t) C | |||||
|---|---|---|---|---|---|---|---|
| A | ke | R2 | B | C | ks | R2 | |
| LL | 187.49 (4.26)bc | 0.11 (0.01)bc | 0.955 | 9.37 (1.01)c | 89.95 (1.05)a | 0.007 (0.002)d | 0.920 |
| VD | 200.81 (4.01)b | 0.23 (0.03)ab | 0.958 | 12.02 (0.82)bc | 86.58 (0.52)a | 0.048 (0.011)bc | 0.911 |
| VP | 165.67 (3.73)d | 0.15 (0.02)abc | 0.954 | 9.95 (1.16)c | 88.82 (0.61)a | 0.119 (0.048)ab | 0.787 |
| AS | 171.05 (4.27)cd | 0.16 (0.02)ab | 0.939 | 12.10 (0.70)bc | 86.51a (0.56)a | 0.020 (0.003)bcd | 0.937 |
| SM | 154.63 (3.77)d | 0.08 (0.01)bc | 0.954 | 10.73 (1.98)bc | 87.97 (2.08)a | 0.006 (0.003)d | 0.837 |
| VM | 242.31 (4.66)a | 0.32 (0.04)a | 0.959 | 15.57 (1.02)b | 83.50 (0.47)b | 0.252 (0.053)a | 0.923 |
| XA | 165.99 (3.61)d | 0.17 (0.02)ab | 0.958 | 28.33 (1.32)a | 69.71 (0.88)c | 0.036 (0.006)bc | 0.956 |
| SG | 165.95 (4.50)d | 0.03 (0.004)c | 0.956 | 13.30 (1.33)bc | 85.97 (1.37)ab | 0.008 (0.002)d | 0.919 |
Changes in LMR
The LMR of all species decreased rapidly from 81.1 to 97.6% in the first 24 h respectively in X. americana and L. lanceolata (Fig. 2). For all species, half of the mass loss by leaching occurred within 3 days (72 h). After 15 days of immersion in water, the LMR varied from 66 to 90% of its initial mass, respectively in the same previous species, corresponding to a mass loss between 34 and 10% of their initial mass.
Fig. 2 - Changes in litter mass with leaching time in the 8 agroforestry species studied. AS: A. senegalensis; LL: L. lanceolata; SG: S.g. guineensis; SM: S. g. macrocarpum; VP: V. paradoxa; VD: V. doniana; VM: V. madiensis; XA: X. americana.
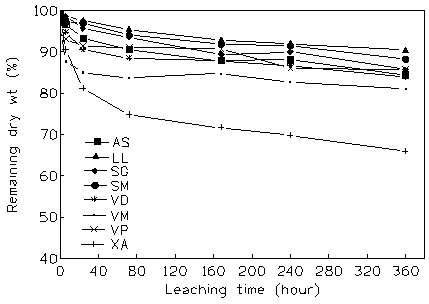
The LMR was fitted to negative exponential model, with a highly significant coefficient of determination for all species (Tab. 2). A multiple comparison of leaching rate constants (ks ) and water soluble compartment (B) by T’-method ([28]) showed that the changes in mass loss during the leaching experiment varied among species. V. madiensis had the highest leaching rate constant and L. lanceolata and both Syzygium species the lowest ones. Nevertheless, X. americana distinguished itself by the highest water soluble substance compartment and L. lanceolata and V. paradoxa by the lowest ones.
Changes in organic compounds
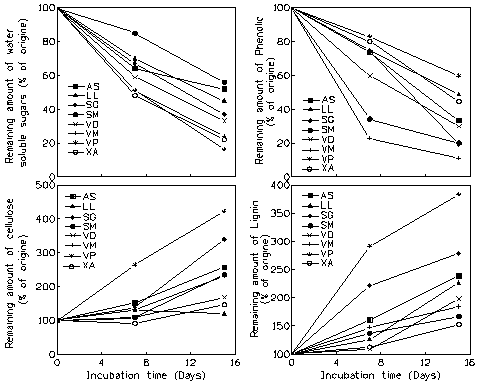
The chemical composition (%) of litter generally changed by differential losses or gains of its compounds during 360 h of leaching (Tab. 1). However, in some species no change was observed in water-soluble sugars (S. g. macrocarpum), cellulose (L. lanceolata), phenol (L. lanceolata and V. paradoxa) and lignin (S.g. macrocarpum and V. madiensis). Organic compounds changes after 168 and 360 h of litter leaching were presented in Fig. 3. The quantity of water-soluble sugars and phenol compounds decreased significantly during leaching in seven species (Fig. 3 and Tab. 3): 48 -83% of water-soluble sugars and 40 -89% of phenol were leached after 15 days of immersion in water, i.e. 0.21 - 0.50 g of sugars and 0.06 - 0.05 g of phenol, respectively in A. senegalensis and V. paradoxa for sugars and in V. paradoxa and V. madiensis for phenol. In contrast, cellulose and lignin increased during the same process in the all species, with differences not always significant (Tab. 3): 46 - 322% of cellulose and 98 - 283% of lignin were gained after 15 days of immersion in water, i.e. 0.07 - 0.22 g of cellulose, respectively in X. americana and V. paradoxa, and 0.15 - 0.14 g of lignin respectively, in V. doniana and V. paradoxa.
Fig. 3 - Changes in litter organic compounds (water soluble sugars, phenol, cellulose and lignin) with leaching time in the 8 agroforestry species studied. AS: A. senegalensis; LL: L. lanceolata; SG: S.g. guineensis; SM: S. g. macrocarpum; VP: V. paradoxa; VD: V. doniana; VM: V. madiensis; XA: X. americana.
Tab. 3 - Mean differences in organic compound amounts between initial and final litter (360 h of incubation) expressed in g and, if these differences were significant, also as percentage of the initial amount. ND: non determined; ns: non significant; * P < 0.05; ** P < 0.01; *** P < 0.001. Gain () and loss (-). AS: A. senegalensis; LL: L. lanceolata; SG: Syzygium guineense var. guineense; SM: S. guineens e var. macrocarpum; VD: V. doniana; VM: V. madiensis; VP: V. paradoxa and XA: X. americana.
| Parameter | Deciduous broad-leaved trees | Deciduous broad-leaved shrubs | Evergreen broad-leaved tree | ||||||
|---|---|---|---|---|---|---|---|---|---|
| LL | VD | VP | AS | SM | VM | XA | SG | ||
| Litter mass | %g | -9.61*** | -16.09*** | -14.17*** | -15.56*** | -11.8*** | -18.89*** | -34.12*** | -14.27*** |
| Soluble sugars | G | -0.20* | -0.33** | -0.50** | -0.21*** | -0.12ns | -0.40** | -0.41* | -0.20* |
| % | 54.05 | 66.84 | 83.24 | 47.98 | ND | 75.44 | 77.40 | 65.58 | |
| Phenol | G | -0.04ns | -0.05** | -0.06* | -0.06** | -0.14** | -0.05*** | -0.08** | -0.12** |
| % | ND | 70.23 | 40.40 | 66.60 | 80.05 | 89.19 | 55.36 | 81.00 | |
| Cellulose | G | 0.02ns | 0.10* | 0.22*** | 0.15*** | 0.11** | 0.13*** | 0.07* | 0.18*** |
| % | ND | 67.00 | 321.65 | 156.86 | 134.97 | 130.21 | 45.94 | 239.60 | |
| Lignin | G | 0.33* | 0.15* | 0.14* | 0.23* | 0.10ns | 0.09ns | 0.08ns | 0.22** |
| % | 126.60 | 97.93 | 283.10 | 138.50 | ND | ND | ND | 178.9 | |
Correlations between water absorption and leaching rate constants and initial litter properties
Tab. 4 - Correlation coefficients of Pearson (n = 8) calculated between initial litter properties and maximum water content (MWC), maximum water soluble substances (MSS) and water absorption and leaching rate constants. In bold significant (P < 0.05 or P < 0.01) correlations.
| Parameter | MWC | ke | MSS | ks |
|---|---|---|---|---|
| Thickness (mm) | 0.2095 | 0.1923 | -0.3767 | -0.0722 |
| Area (cm2) | -0.3876 | 0.9420 | -0.7858 | -0.3679 |
| Initial WC (%) | -0.4179 | -0.5331 | -0.6753 | -0.1889 |
| Water-soluble sugars (%) | 0.7891 | 0.9420 | 0.3752 | 0.8611 |
| Phenol (%) | -0.8245 | -0.7173 | 0.1973 | -0.3932 |
| Cellulose %) | -0.0066 | 0.4251 | 0.4152 | 0.3518 |
| Lignin (%) | -0.0407 | -0.3531 | -0.1162 | -0.6021 |
The asymptote of the absorption exponential model (or maximum water content = MWC) was correlated positively with water-soluble sugar contents and negatively with phenol contents of initial litter (Tab. 4). The water absorption rate constant (ke) was positively correlated with initial litter area and water-soluble sugars and negatively correlated with phenol content of initial litter. The compartment of water-soluble substance (MSS) was negatively correlated with litter area, while the leaching rate constant (ks) was positively correlated with water-soluble sugars. It was also correlated with maximum water absorption capacity (or maximum water content = MWC) of litter (eqn. 3):
The water content of litter at the end of the leaching experiment was not correlated with the total mass loss. Nevertheless there was significant correlation between the absorption rate constant (ke ) and the leaching rate constant (ks) of the litter (R2= 0.8011, P < 0.005, n = 8). The stepwise model showed that the relationship between water absorption rate constant (ke) and initial litter properties was explained about 90% by the association of litter thickness and water-soluble sugars (eqn. 4):
The relationship between leaching rate constant (ks ) and initial litter properties was explained about 95% by the association of water-soluble sugars and initial water content of initial litter (eqn. 5):
Discussion and conclusions
Water absorption and mass loss were rapid at the start of incubation for all species. The dynamics of each of the both processes slowed until 72 h when the water absorption and the litter mass loss reached their maximum. No data from litter of African tropical savannas was available in literature, excepting that of Ibrahima et al. ([15]). Their study found that the water content of nine species of sudano-guinea savanna of Adamawa, including the eight species under present study varied from 104 to 240% of dry mass respectively for S. g. var. guineense and Uapaca togoensis during 24 h of leaching in laboratory, and from 173 to 335% for the same species at the end of experiment (240 h). The mass loss varied between 5 and 28% of initial dry mass respectively for V. paradoxa and X. americana during 24 h of leaching and from 12 and 33% for the same species during 240 h of leaching. In the present study, the litter water content varied from 88.21% (S. g. var. guineense) to 226.90% (V. madiensis), in 24 h, from 158.66 (S. g. var. macrocarpum) to 245.85% (V. madiensis), in 240 h, and from 162.77% to 264.52% of their dry mass in the same species at the end of the experiment (after 360 h). The mass loss varied from 2.39% (L. lanceolata) to 18.87 (X. americana), in 24 h, from 8.00 to 30.17%, in 240 h, and from 9.61 to 34.12% in the same species at the end of the experiment (360 h). The difference between the two results were significant (P < 0.05) for eight species (not including U. togoensis) during 240h of leaching, despite the two occurred in the different period.
Similar results were reported by Nykvist ([22], [23], [24]), Taylor & Parkinson ([30]), Parsons et al. ([26]) and Ibrahima et al. ([14]) on the leaf litter leaching of Temperate and Mediterranean broad-leaved species. According to Taylor & Parkinson ([30]), the water absorption and the litter mass loss of trembling aspen (Populus tremuloides), in 24 h, were respectively 300% of litter dry mass (either about 90% of maximum water absorption capacity) and 24% of initial dry mass. Ibrahima et al. ([14]) also have found that the litter of six Mediterranean tree species have the water absorption capacity ranging from 98%, in Quercus ilex, to 250%, in Castanea sativa, in 24 h of incubation, reaching, in 240 h of incubation, 155% and 362% of dry mass, respectively. The mass loss varied from 3.2%, in Fagus sylvatica, to 6% of their initial mass, in Castanea sativa, during 24 h of incubation, and from 7% to 15% in the same species during 240 h of incubation.
Leaf Litter of Sudano-guinea Savannas of Ngaoundere seemed to have different patterns of water absorption and leaching than those of Temperate and Mediterranean regions. The Temperate leaf litters (Castanea sativa and Populus tremuloides) have high water absorption capacity and high absorption and leaching rate constants. Water absorption and mass loss were almost completed (reached 90% of water absorption capacity) in 24 h of leaching in Populus tremuloides, Fraxinus excelci or and Castanea sativa ([24], [30], [14]), while in the Mediterranean and Sudano -guinea Savannas species, water absorption and mass loss slowed down and reached their maximum capacity around 72 h, even 240 h of incubation time varying with the species. Nevertheless, water -soluble substances (or maximum mass loss) of Sudano -guinea Savannas leaf litters (31% of initial dry mass in 240 h) were about 1.3 and 2 times higher than those of the Temperate (24%) and Mediterranean (15%) species, respectively. These results were in conformity with those reported by Jung et al. ([16]) in their studies on the comparison among some litters of Temperate species (Calluna vulgaris, Festuca sylvatica and Fagus sylvatica) and Tropical species (Acacia albida, Guiera senegalensis, Khaya senegalensis and Gmelina arborea) of the dry Sahel-Senegalian Savannas. They have shown that the litters of tropical species were generally richer in water-soluble substances than those of temperate species. Other studies ([4], [5]) had shown that Eucalyptus and Acacia auriculiformi leaf litters, originating from the Australian region and now largely planted in the African Savannas, exhibited a high content of soluble organic matter, ranging from 30% to 35% of dry mass. This suggested that the leaching role (or initial phase of litter decomposition) possibly has an important implication in the litter decomposition processes of Sudano -guinea Savannas, insofar as it is in this stage that energetic substances, such as water-soluble sugars, necessary for the biological activities, were mostly released.
According to the study of Ibrahima et al. ([14]), the litters of deciduous species as Castanea sativa and Fagus sylvatica were distinguished from these of evergreen species like Quercus ilex and Quercus coccifera by their higher water absorption and higher mass loss during 10 days of laboratory leaching experiment in darkness. Conversely, no net differences were revealed in this study, among evergreen (S. g. guineense) and deciduous species (all remaining species), among forestry gallery species (S. g. guineense, V. doniana) and dry upland Savanna species (A. senegalensis, L. lanceolata, S. g. macrocarpum, V. madiensis, V. paradoxa and X. americana), nor among tree species (L. lanceolata, S. g. guineense, V. doniana and V. paradoxa) and shrubs ones (A. senegalensis, S. g. macrocarpum, V. madiensis, X. americana) according to water absorption capacity and water-soluble substance loss by their litters during 360 h of leaching. But, differences among plant families and among plant species seemed to appear in the present study. In fact, water absorption and release of water-soluble substances rate constants of Myrtaceae (S. g. guineense and S. g. macrocarpum) were slower than those of Verbenaceae (V. doniana and V. madiensis). In the same way, leaf litter of X. americana was richer in water-soluble substances than those of other species, while water absorption rate constant showed the highest value in V. madiensis and the lowest one in S. g. guineense, L. lanceolata and S. g. macrocarpum. This difference among species was reported by Jung et al. ([16]) who studied the quality (but not the quantity) of water-soluble substances in the various types of litter. They thus showed that the litter of Acacia albida is richer in reducing sugars and contains relatively low amounts of amino -nitrogen than Khaya senegalensis and Gmelina senegalensis. They demonstrated that the supplementary contribution of Acacia leaves entails a marked enriching in all the water-soluble compounds of a little-leached tropical ferruginous soil. According to these authors, this fact partially explains the favourable influence of Acacia on the biological activities of the soil, proven in previous works ([17]) and on the output of crops, reported by Charreau & Vidal ([9]).
Generally the physical features of leaf-litter as well as their chemical quality seemed to play a great role in the changes in water absorption and water soluble substances, as reported by Taylor & Parkinson ([30]), Saini ([27]) and Ibrahima et al. ([14]). The maximum water absorption (A) and its rate constant (ke ) were positively influenced by litter area and thickness and water-soluble sugars, and negatively affected by initial phenol content, while litter mass loss (MSS) and their rate constants (ks ) were negatively influenced by litter area and positively affected by water soluble sugars and initial water content of litter. During the 15 days of immersion in water, while at the same time absorbed water, the litter lost mass by releasing some of its constituents, which became dissolved in the water. But, no relation was found between the absolute value of water absorption by litters and their mass loss during leaching. This relationship exists between the rate constants of the two processes. These results did not agree with the findings of Ibrahima et al. ([14]) on the Mediterranean litter types, because the authors reported that the initial cellulose content of litter seemed to play a role in the rate at which the litters absorbed water and the leaf thickness did not seem to have any influence on the dynamics of mass loss during leaching. On the other hand, in our study, litter thickness when it is combined with water-soluble sugars, seemed to influence the rate at which the litters absorbed water. Initial cellulose content did not play any role in the two processes in our study. Similarly, the quality index of initial lignin content of litters often considered as a determining factor for the overall speed of decomposition (e.g., [31], [7], [12]), did not seem to influence the dynamics of this leaching phase of decomposition. This is probably because decomposition mainly results from both physical, chemical and biological processes, but the leaching phase results only from physical and chemical processes ([14]).
During the 15 days of leaching, the loss of dry mass was on average 9-34 g for each 100 g of initial litter. Among the C-rich constituents, soluble sugars and phenol were leached to a large extent: 48 - 83% and 40 - 89% of the sugars and phenol initially present in the litter disappeared during the 15 days of the experiment. This loss seemed to be proportional to the initial compound content of the litter. For instance, V. paradoxa, whose litter was the richest in soluble sugars and the poorest in cellulose and lignin content, had the lowest amounts in soluble sugars after 15 days after leaching. Comparatively, the quantity of lignin and cellulose in the litter increased during the 15 days of incubation. Metabolic compounds, which account for most of the constituents released by leaching, are molecules that are readily metabolized by micro-organisms; it is known that micro-organisms capable of degrading lignin compounds needed also other C sources ([3]). Leaching of fresh litter can therefore lead to an export of these labile compounds towards the soil and therefore provide a supply of metabolic compounds for micro-organisms that participate in the decomposition processes of the underlying older litter layers. The mineral elements released by leaching are probably quickly recycled in the soil and vegetation.
Leaching changed the chemical composition of the litters by lowering the water-soluble sugar concentration (significant in 7 species) and phenol concentration (significant in 6 species), and increasing the cellulose (significant in 7 species) and lignin (significant in 6 species) concentrations. These results agree with those of the literature. According to our results and those of Parsons et al. ([26]) and Ibrahima et al. ([14]), there is therefore a consistent trend in litter quality during leaching. Leaching may therefore have more important consequences for the subsequent decomposition processes in the Sudano-guinea Savannas of Ngaoundere, as well as in the tropical Savannas ([5]), since it is known that the initial content of water-soluble substances plays a major role during litter decomposition. Berg & Tamm ([2]) and Berg & Ekbohm ([1]) showed that there is in fact a positive relation between the initial quantities of water-soluble substances in the litter and its rate of decomposition. Similarly, Mangenot & Toutain ([20]) demonstrated that the litter types that are richest in water-soluble substances are also those that are most quickly decomposed. We have also shown ([13]) that the dynamics of mass loss during the decomposition of 12 Mediterranean litters in microcosms was largely explained by their initial content of water-soluble substances. Prolonged leaching could therefore transform fresh litter into a material that is less readily biodegradable. Parsons et al. ([26]) demonstrated this. They showed that P tremuloides litter that had been leached for 5 days subsequently decomposed much more slowly than intact litter. They also showed that the rate of mass loss from leaching litter was constant with time, whereas intact litter followed an exponential decline. According to the authors, this was probably due to the loss of a great proportion of the labile substances in the litter and therefore to the suppression at this stage of rapid mass loss at the start of decomposition.
In conclusion, the litters of the Sudano-guinea Savannas of Ngaoundere used in our study showed different capacities to release water-soluble substances during leaching. Three groups were distinguished: X. americana with the highest capacity to release water-soluble substances (34.12%), in contrast, L. lanceolata and both Syzigiums showed the lowest capacity to release water-soluble substances (9.61 to 14.27%) and A. senegalensis and both Vitex (15.56 to 18.89%) had intermediate patterns. The leaching rate constant of these litters was influenced by physical and chemical characteristics of original litters as reported in Temperate and Mediterranean biomes. Due to the high capacity to release water -soluble substances, such as water-soluble sugars, most of the Sudano -guinea Savannas litters have to be integrated into the farming systems to replace exotic plants species, as eucalyptus, whose water-soluble substances play a negative role in plant decomposition by slowing down the litter decomposition rate constant ([5], [6]).
Acknowledgements
The research was supported partly by the University of Ngaoundere through of GPRU and partly by the International Foundation for Science (IFS), Stockholm, Sweden and United Nation University (UNU), Tokyo, Japan, through a grant to M. Adamou Ibrahima.
References
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
M Halima
Department of Biological Sciences, Faculty of Science, The University of Ngaoundere, P.O.Box 454, Ngaoundere (Cameroon)
Department of Food and Nutritional Sciences, National High School of Agro-Industrial Sciences, The University of Ngaoundere, P.O.Box 455, Ngaoundere (Cameroon)
Corresponding author
Paper Info
Citation
Ibrahima A, Biyanzi P, Halima M (2008). Changes in organic compounds during leaf litter leaching: laboratory experiment on eight plant species of the Sudano-guinea Savannas of Ngaoundere, Cameroon. iForest 1: 27-33. - doi: 10.3832/ifor0450-0010027
Paper history
Received: May 26, 2007
Accepted: Oct 10, 2007
First online: Feb 28, 2008
Publication Date: Feb 28, 2008
Publication Time: 4.70 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2008
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 60265
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 47023
Abstract Page Views: 4730
PDF Downloads: 7202
Citation/Reference Downloads: 62
XML Downloads: 1248
Web Metrics
Days since publication: 6713
Overall contacts: 60265
Avg. contacts per week: 62.84
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2008): 15
Average cites per year: 0.83
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Short Communications
Effects of nitrogen, calcium and cation exchange capacity on gum yield in Acacia senegal under plantation and savanna woodland conditions in northern Guinea savanna, Nigeria
vol. 4, pp. 190-194 (online: 11 August 2011)
Research Articles
Soil fauna communities and microbial activities response to litter and soil properties under degraded and restored forests of Hyrcania
vol. 14, pp. 490-498 (online: 11 November 2021)
Research Articles
Carbon, nitrogen and phosphorus stoichiometry controls interspecific patterns of leaf litter-derived dissolved organic matter biodegradation in subtropical plantations of China
vol. 14, pp. 80-85 (online: 19 February 2021)
Research Articles
Litter quality changes during decomposition investigated by thermal analysis
vol. 8, pp. 827-837 (online: 19 March 2015)
Research Articles
The manipulation of aboveground litter input affects soil CO2 efflux in a subtropical liquidambar forest in China
vol. 12, pp. 181-186 (online: 10 April 2019)
Research Articles
Effects of stand age on litter quality, decomposition rate and nutrient release of Kazdagi fir (Abies nordmanniana subsp. equi-trojani)
vol. 13, pp. 396-403 (online: 03 September 2020)
Research Articles
Impact of deforestation on the soil physical and chemical attributes, and humic fraction of organic matter in dry environments in Brazil
vol. 15, pp. 465-475 (online: 18 November 2022)
Research Articles
Fine root production and distribution in the tropical rainforests of south-western Cameroon: effects of soil type and selective logging
vol. 3, pp. 130-136 (online: 27 September 2010)
Research Articles
Nutrients (N, P and K) dynamics associated with the leaf litter of two agroforestry tree species of Bangladesh
vol. 2, pp. 183-186 (online: 15 October 2009)
Research Articles
Litter quality and decomposition rates of selected species in the Ankasa Conservation Area, Ghana
vol. 19, pp. 122-129 (online: 11 April 2026)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







