
Combined pre-hardening and fall fertilization facilitates N storage and field performance of Pinus tabulaeformis seedlings
iForest - Biogeosciences and Forestry, Volume 9, Issue 3, Pages 483-489 (2016)
doi: https://doi.org/10.3832/ifor1708-008
Published: Jan 07, 2016 - Copyright © 2016 SISEF
Research Articles
Abstract
Exponential fertilization during the pre-hardening stage and fall fertilization during the hardening stage have each been used independently to nutrient load seedlings. However, nursery and field responses of seedlings to the combination of exponential fertilization and fall fertilization have received little attention. Chinese pine (Pinus tabulaeformis Car.) container seedlings were exponentially fertilized with accumulated totals of 40, 80 or 120 mg N per seedling during pre-hardening, and fall-fertilized with 0, 12, 24 or 48 mg N per seedling, and were subsequently outplanted and followed for two growing seasons. Interactions of exponential and fall fertilization had significant effects on plant N content in the nursery and first-year height after outplanting. Fall fertilization promoted additional nutrient loading during hardening for the 40-80 mg N per seedling pre-hardening regimes. The highest exponential fertilization rate enhanced N concentration in foliage and roots compared to the other two rates. Maximum diameter was observed in the lowest exponential fertilization rate at the second year after outplanting. Fall fertilization enhanced foliar N concentration. Supplemental 12 and 24 mg N per seedling during fall were more effective in increasing height increment at the second year after outplanting. Our results indicate that pre-hardening fertilization is a useful tool to nutrient load Chinese pine in the nursery and facilitate outplanting performance in the field. In combination, fall fertilization has potential to further augment this response, although further research is needed to precisely match rates of pre-hardening and fall fertilization to optimize seedling performance.
Keywords
Exponential Fertilization, Fall Fertilization, Nutrient-loading, Field Performance, Chinese Pine
Introduction
More nitrogen (N) is retranslocated from perennial tissues to new apical growth tissues in seedlings with rich N reserves, especially when seedlings are outplanted in poor fertility soil ([34], [27], [48]). Thus, additional N fertilization during nursery culture may provide an effective solution for enhancing field performance and has received worldwide attention ([40], [1], [48], [29]).
Exponential fertilization during the pre-hardening stage and fall fertilization during the hardening stage are two approaches to facilitate N storage in seedlings ([40], [39], [9]). Exponential fertilization reflects application of fertilizer at exponential rates corresponding closer to the desired relative growth rate of plants during their exponential phase of growth ([40], [3]). It synchronizes nutrient supply with crop demand, referred to as steady-state nutrition ([15]), and has proven to facilitate nutrient storage in a variety of tree species ([41], [26], [31], [33], [35], [30]). Previous studies comparing one exponential fertilization rate with one or more conventional fertilization rates, have effectively demonstrated that greater plant N reserves induced by exponential fertilization have the potential to promote seedling field performance over conventional fertilization ([10], [6], [51]). When outplanted onto a field site, how seedlings respond to varying exponential fertilization rates in the nursery has been studied on several species, including western hemlock (Tsuga heterophylla (Raf.) Sarg. - [12]), northern red oak (Quercus rubra L.) and white oak (Q. alba L. - [37]). However, less attention has been given to pines (Pinus spp.), which are among the most widely distributed and planted species worldwide.
Fall fertilization is another approach to facilitate nutrient loading. Fall fertilization could counter nutrient dilution due to biomass accumulation and promote storage of nutrient reserves in seedlings ([45]). Therefore, it has been implemented in a variety of tree species such as loblolly pine (Pinus taeda L. - [39]), slash pine (Pinus elliottii var. elliottii Engelm. - [16]), red pine (Pinus resinosa Ait. - [17]), Norway spruce (P. abies (L.) Karsten - [32], [13]), black spruce (Picea mariana Milli.B.S.P. - [4]), Lutz spruce (Picea x lutzii Littl. - [19]), Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco - [45], [23], [24]), holm oak (Q. ilex spp. ballota Desf. - [28], [1]), Olga Bay larch (Larix olgensis Henry - [20], [54]).
In addition, the efficacy of fall fertilization on seedling nutrient reserves is dependent on pre-hardening nutrient status ([4], [21]). If seedlings are provided with high rates of fertilizer and thus have substantial amounts of stored nutrients during the pre-hardening period, then fall fertilization has little additional impact on nutrient reserves ([4]). For example, in the deciduous Chinese cork oak (Quercus variabilis Blume), root N concentration was affected by the interaction of pre-hardening and fall fertilization ([21]). Due to variable strategies of nursery fertilization among tree species ([44]), little information is available on how the combination of pre-hardening and fall fertilization impacts nutrient storage of Pinus spp. seedlings.
Stored nutrients in nursery seedlings are often correlated with early seedling survival after outplanting ([45], [48]), and nutrient differences among fall fertilization treatments are influenced by pre-hardening fertilization ([4], [21]). Earlier papers have primarily dealt with the relationship between field performance and pre-hardening or fall fertilization independently ([31], [32], [12], [6], [10], [28], [29]), but expanded study aimed at understanding how their interactive effects impact nursery and field performance are needed, especially for species of widely planted genera such as Pinus.
Chinese pine (Pinus tabulaeformis Carr.) is a native evergreen coniferous species and the most widely planted species in northern China. Because of its resistance to drought and low soil fertility, it is planted across a wide range of site types and is often exposed to environmental stresses ([53], [52]). Nursery fertilization has been shown to promote Chinese pine seedling nutrient reserves ([14], [50]), suggesting potential to increase stress resistance and improve outplanting performance ([48], [29]). Thus, our objectives were to: (1) examine the combined effects of these two nutrient loading approaches on Chinese pine seedling nutrient storage in the nursery; and (2) assess how field performance responds to the application of combined exponential and fall fertilizations in the nursery. We focused on N because this nutrient is often the most limiting factor for seedling growth in peat-growing media ([11]), is required in the largest amount for intensive nursery culture of conifers ([35]), and is highly correlated with field performance after outplanting ([48], [29]). We hypothesized that: (1) pre-hardening and fall fertilization would each promote nutrient reserves in Chinese pine seedling yet vary in their effectiveness; (2) pre-hardening and fall fertilization would be more advantageous in combination than individually; and (3) outplanting performance would be strongly correlated with seedling growth and nutrient storage from the nursery phase.
Materials and methods
Nursery fertilization treatments and outplanting trial
The experiment was conducted in a greenhouse at the Chinese Academy of Forestry Sciences in Beijing (40° 40′ N, 116° 14′ E). Our experiment investigated the main and interacting effects of three rates of pre-hardening N fertilization and four rates of N fertilization during hardening (fall fertilization). On 21 February 2010, seeds from a regional seed orchard (National Seed Orchard for Chinese Pine, Qigou Forest Farm, 41° 00′ N, 118° 27′ E, 526 m a.s.l.) were sown into plastic containers (8 cm diameter × 8 cm deep, 187 ml capacity) filled with a 3:1 (v:v) peat:vermiculite medium. Thirty containers were randomly assigned to a tray (53 cm long × 44.5 cm wide), providing a density of 127 seedlings m-2.
Three pre-hardening fertilization rates based on another Pinus species study ([26]), starting 1 week after germination and continuing for 25 weeks, were tested: (i) 40 mg N per seedling; (ii) 80 mg N per seedling; and (iii) 120 mg N per seedling following exponential (E) functions ([25], [10]). A total of 48 trays were arranged in a completely randomized design with sixteen trays for each rate.
In this study, the average initial N content per seed (NS) was determined to be 1.58 mg N. The total number of fertilizations (t) was 25. N was supplied as urea (Xilong Chemical Co., China), a N source commonly used in Chinese nurseries. Elemental P and K were supplied as KH2PO4 (Guangdong Guanghua Sci-Tech Co. Ltd., China). On 16 August the pre-hardening fertilization was terminated, each seedling received total amounts of 26.2 mg P and 32.9 mg K which were evenly split and applied weekly in addition to the N fertilizer (1 March-16 August, 2010). The desired amounts of N, P and K were dissolved in distilled water and added manually to each seedling once per week with a hand-sprayer. No micronutrients were applied during the experiment. Seedlings were rinsed after each application to avoid foliar fertilizer burn. Additional irrigation was provided as necessary, about two times each week. The trays on raised benches were rotated every week to minimize edge effect.
Fall fertilization started two weeks after the pre-hardening fertilization ended when most seedlings had already formed the terminal buds, and lasted for four weeks (30 August-20 September, 2010). The doses of N were evenly split and applied weekly as four fall fertilization rates: (i) control (no additional fertilization); (ii) 12 mg N per seedling; (iii) 24 mg N per seedling; and (iv) 48 mg N per seedling. Fall fertilization was applied within each of the three pre-hardening regimes (40, 80, 120 mg N per seedling) as a factorial 3×4 design that was randomly arranged with four replications per treatments combination. The desired amount of N was supplied as urea; 20 ml of appropriate fertigation solution was added by hand to each seedling. On 24 October, all seedlings were moved outdoors to hasten hardening. After 28 November, seedlings were stored under snow cover over winter.
Temperature was measured with a JL-18 Series thermometer (Huayan Instrument and Equipment Co., Shanghai, China) at 10-min intervals. From sowing throughout the end of the pre-hardening fertilization, the daily average for day:night temperature in the greenhouse was 25:18 °C, respectively. During the fall N application period and until the seedlings were moved outdoors, greenhouse temperatures averaged 19:14 °C (day:night). Once outside, seedlings were exposed to temperatures averaging 13:9 °C (day: night).
On 1 May 2011, seedlings from the twelve fertility treatments were shipped and outplanted in a previously cultivated land at the Beijing Forestry University Northern Experimental Base at Pingquan, Hebei province (41° 13′ N, 118° 40′ E). The soil was ploughed before outplanting. Weeds were removed by hand. The site has an average elevation of 765 m a.s.l. with a small slope (< 2%). The depth of soil varied between 45 and 60 cm. The surface soil (0-20 cm) is 73% sand, 11% silt, and 16% clay (a sandy clay loam) with a pH of 6.2 and soil organic carbon of 0.7%. Average total N, available P, and available K were 628.7, 139.5, and 113.5 mg kg-1, respectively. The soil was considered to have moderate fertility according to the macronutrient classification criteria ([2]). The area has a temperate continental monsoon climate, characterized by dry winter and spring seasons. During 2011 and 2012, mean annual air temperature was 7.4 °C and 6.8 °C, respectively. Annual rainfall was 486 mm in 2011 and 626 mm in 2012.
The field experiment was arranged as a randomized complete block experimental design with four replicates. Each block measured 14×16.5 m2 and was separated from adjacent blocks by 1 m buffers. Fifteen randomly selected seedlings from a tray per treatment were planted in single parallel rows within each block, resulting in a total of 720 seedlings planted. Seedlings were planted in 0.4×0.4×0.4 m manually dug planting holes with a 1×1.5 m2 spacing.
Harvesting and measurements
Seedlings were sampled prior to planting to evaluate biomass allocation and N storage responses to nursery fertilization. Thirty-two seedlings per nursery fertilization treatment (eight seedlings per replicated tray) were randomly chosen. After carefully washing, seedlings were separated into needles, stems and roots. Organs of seedlings were oven-dried at 65 °C for 48 h to determine biomass. Organ N concentration was determined from a composite sample of eight seedlings for each treatment from a tray using semi-micro Kjeldahl distillation ([5]).
Immediately after planting on 2 May 2011, seedling height and diameter were measured (T1). When seedling growth ceased in late October of 2011 (T2) and 2012 (T3), survival, height and diameter were measured. Survival was calculated as the number of saplings of the original fifteen remaining alive for each nursery-treatment replicate at the time of each annual measurement. Diameter was measured at ground rate on each tree stem, and total seedling height was measured from the root collar to the tip of the terminal bud. Net increment of height in cm (or diameter in mm) was defined as the difference observed between T2 and T1, and between T3 and T2.
Statistical analyses
Effects of pre-hardening fertilization (3 rates), fall fertilization (4 rates), and their interactions were assessed using a two-way analysis of variance (ANOVA) applied on a completely randomized design at the nursery experiment and on a randomized complete block design in the outplanting trial. When ANOVA results showed a significant effect, post-hoc Duncan’s test was carried out for multiple comparisons among treatments (α=0.05). Statistical analyses were performed using SPSS® 16.0 (Chicago, Illinois, USA). The “explore” function of SPSS was used to examine data prior to ANOVA, and survival was arsine-transformed to fulfill normality and variance homogeneity requirements.
Results
Nursery phase
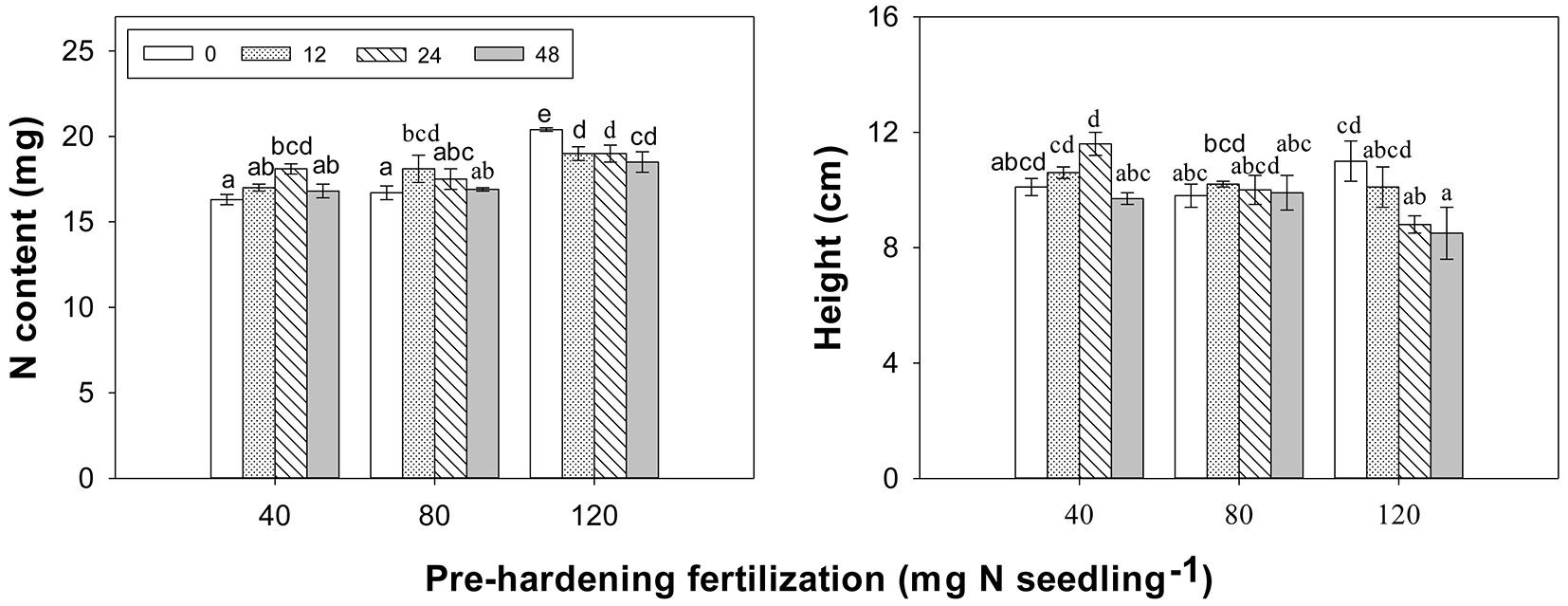
Fertilization factors and its interaction did not significantly affect biomass of specific organs or the whole plant (Tab. 1). The foliage, stem, and root biomass of seedlings averaged 0.33 ± 0.01, 0.16 ± 0.001, and 0.40 ± 0.01 g, respectively. N concentration in foliage, root and whole plant was higher at 120 than at 40 and 80 mg N per seedling of per-hardening fertilization. Fall fertilization enhanced foliar N concentration relative to the control (Tab. 1, Fig. 1). Organ N content did not differ among pre-hardening fertilization or fall fertilization, although pre-hardening fertilization marginally affected N content of three organs (Tab. 1). Overall mean of N content was 7.7 ± 0.16, 3.5 ± 0.08, and 6.7 ± 0.13 mg in foliage, stem and root, respectively. The N content of whole plants, however, was influenced by the interaction of pre-hardening fertilization and fall fertilization (Tab. 1, Fig. 2). At the lowest pre-hardening fertilization regime (40 mg N per seedling), 24 mg N per seedling of fall fertilization significantly increased the N content. At 80 mg N per seedling of pre-hardening fertilization regime, 12 mg N per seedling of fall fertilization yielded a similar effect.
Tab. 1 - P-values derived from ANOVA of the main effects pre-hardening fertilization (PF), fall fertilization (FF) and of their interaction effect (PF×FF) on Chinese pine seedling biomass, N concentration and N content prior to plating.
| Source | df | Biomass | N concentration | N content | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Foliage | Stem | Root | Plant | Foliage | Stem | Root | Plant | Foliage | Stem | Root | Plant | ||
| PF | 2 | 0.829 | 0.417 | 0.360 | 0.273 | <0.001 | 0.191 | <0.001 | <0.001 | 0.070 | 0.076 | 0.061 | <0.001 |
| FF | 3 | 0.152 | 0.784 | 0.892 | 0.624 | 0.001 | 0.434 | 0.120 | 0.867 | 0.373 | 0.493 | 0.248 | 0.137 |
| PF × FF | 6 | 0.407 | 0.205 | 0.113 | 0.346 | 0.147 | 0.219 | 0.832 | 0.919 | 0.520 | 0.375 | 0.083 | 0.010 |
Fig. 1 - Chinese pine seedling organ N concentration (means ± SE) in relation to pre-hardening and fall fertilization prior to planting. No significant interaction between pre-hardening and fall fertilization occurred in N concentration. Bars marked with different letters significantly differed for each organ and the whole plant after Duncan’s test (α = 0.05).
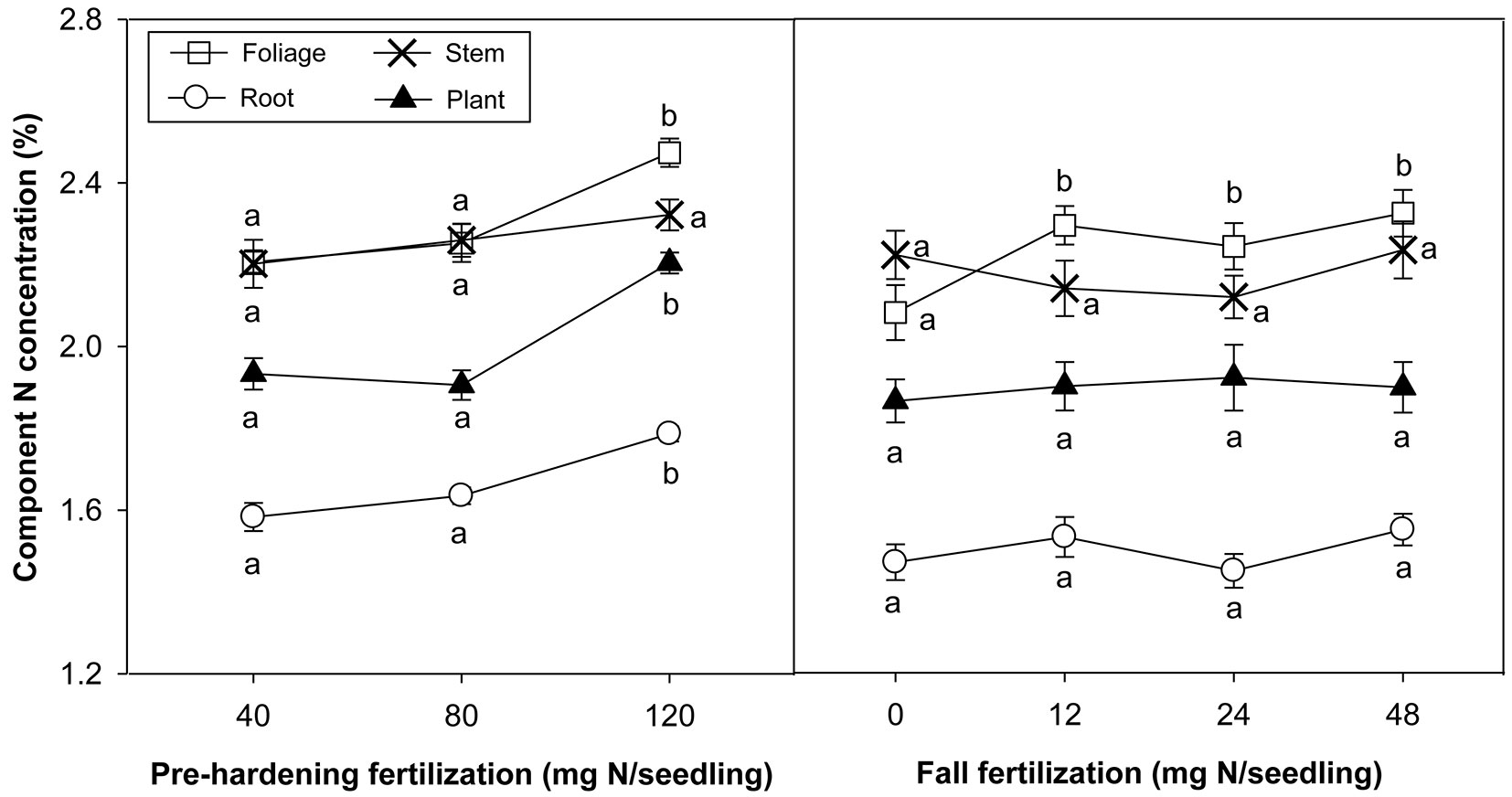
Fig. 2 - Effect of combination treatments between pre-hardening fertilization and fall fertilization (0, 12, 24, 48) on Chinese pine seedling N content (mg N/ seedling) prior to planting and total height (cm) at the end of first growing season (2011) after planting. Means ± SE marked with different letters significantly differed after Duncan’s test (α = 0.05).
Field performance
For all field attributes, the interaction of pre-hardening fertilization and fall fertilization only had a significant effect on total height at the first growing season after outplanting (Tab. 2). The combination of pre-hardening at 40 mg N per seedling and 24 mg N per seedling of fall fertilization (24 mg) yielded the maximum height (Fig. 2). Within the highest pre-hardening fertilization regime, 24 and 48 mg N per seedling of fall fertilization decreased height compared to the control.
Tab. 2 - P-values derived from ANOVA of main effects pre-hardening fertilization (PF), fall fertilization (FF) and of their interaction effect (PF×FF) on Chinese pine seedlings survival, total height, diameter and their increments prior to planting (Initial), at the end of first (2011) and second (2012) growing seasons after planting.
| Source | Height | Height increment | Diameter | Diameter increment | Survival | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Initial | 2011 | 2012 | 2011 | 2012 | Initial | 2011 | 2012 | 2011 | 2012 | 2011 | 2012 | |
| PF | 0.028 | 0.054 | 0.079 | 0.005 | 0.231 | 0.002 | 0.088 | 0.004 | 0.022 | 0.005 | 0.161 | 0.373 |
| FF | 0.458 | 0.087 | 0.031 | 0.021 | 0.029 | 0.303 | 0.201 | 0.309 | 0.138 | 0.366 | 0.458 | 0.793 |
| PF×FF | 0.780 | 0.026 | 0.682 | 0.077 | 0.699 | 0.094 | 0.677 | 0.786 | 0.937 | 0.834 | 0.847 | 0.598 |
Pre-hardening fertilization had no significant effect on survival either during the first or second growing season (Tab. 2). Survival of 40, 80 and 120 mg N per seedling of pre-hardening fertilization was 90%, 95%, and 89% in the first growing season, and was 79%, 79%, and 70% in the second growing season, respectively. Pre-planting height was superior for the highest pre-hardening fertilization rate. However, first-year height increment of later rate was minimum. There was no significant difference in second-year height among pre-hardening rates. The dominance of total height in the highest exponential fertilization vanished at end of the second growing season (Tab. 2, Fig. 3). Both first-year and second-year diameter increments were superior in the 40 mg N per seedling of pre-hardening fertilization treatment, with the greatest diameter occurring in the lowest pre-hardening fertilization rate at the end of second growing season (Tab. 2, Fig. 3).
Fig. 3 - Chinese pine seedlings total height, diameter and their increments in relation to pre-hardening and fall fertilization at planting (Initial), at the end of first and second growing seasons after planting. Error bars represent SE of increments at the end of first and second growing seasons, and of size prior to planting. Bars marked with different capital and lower case letters significantly differed for total height (diameter) and its increment according to Duncan’s test (α = 0.05), respectively. Comparison letters for first-year height are not presented owing to an occurring interaction.
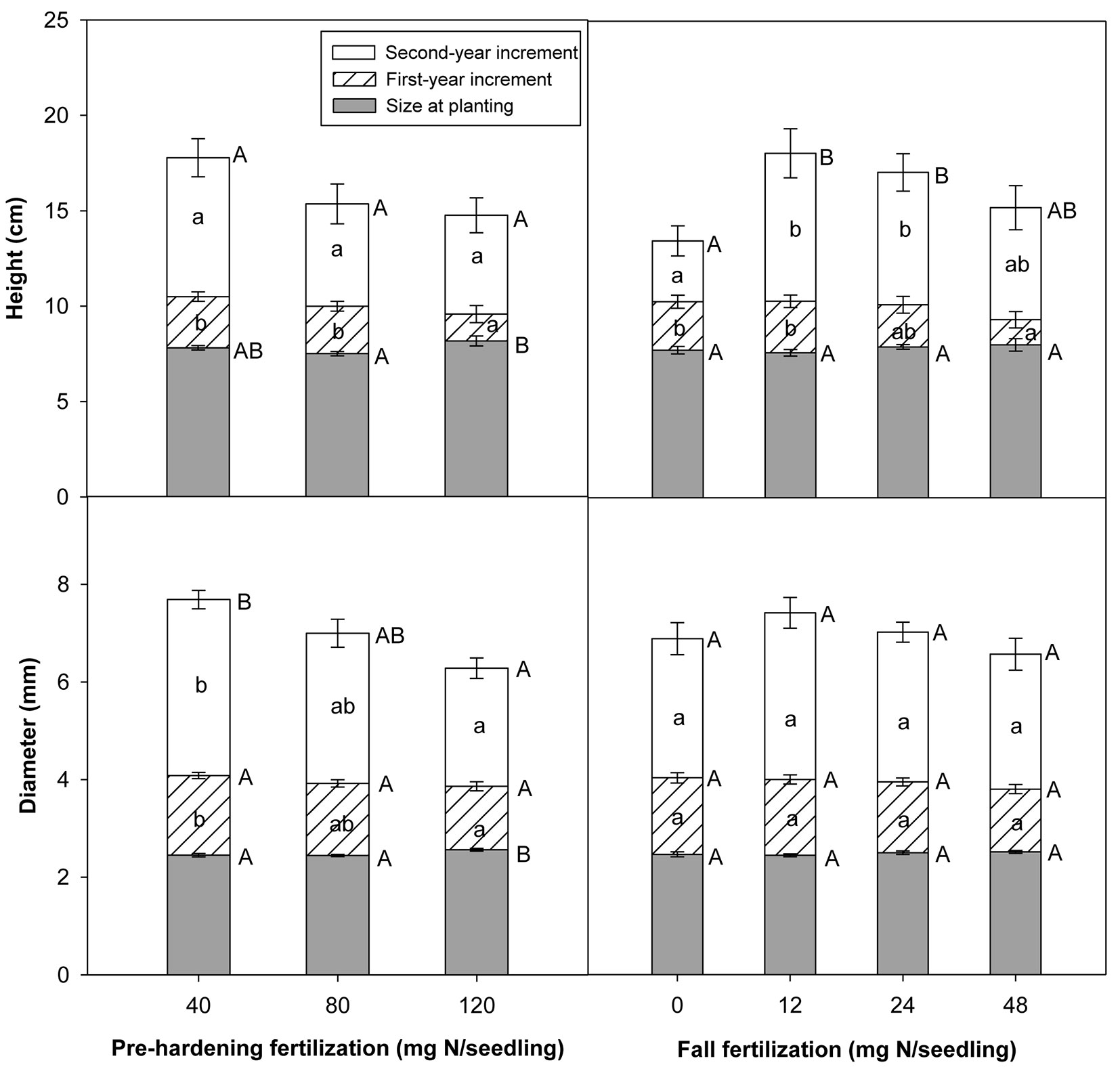
Fall fertilization also did not affect survival of Chinese pine seedlings. Survival of the control, 12, 24 and 48 mg N per seedling of fall fertilization was 92%, 93%, 93%, and 87% in the first growing season, and was 81%, 74%, 77%, and 73% in the second growing season, respectively. Pre-planting height and diameter were similar between fall-unfertilized and fertilized seedlings. Compared to the control, significant height increments in 12 and 24 mg N per seedling of fall fertilization treatments were not observed until the second growing season.
Discussion
Effect of pre-hardening fertilization on nursery and field performance
Under luxury N consumption, seedling dry mass remains unchanged but tissue N content continuously increases ([40], [25], [34], [3]). As defined by the conceptual model ([40]), absence of effects of exponential fertilization on biomass indicated that 40 mg N already reflected luxury consumption for the studied seedlings. This result is similar to our previous study on Chinese pine seedlings ([49]). Deficiency, luxury consumption, and toxicity have been determined in species of Quercus and Picea by evaluating the nursery response to more than six exponential fertilization rates ([35], [3], [36]). For red pine, Timmer & Armstrong ([41]) proposed that 9.75 mg N yielded luxury consumption. In another study of red pine, however, 25 mg N was not designated as luxury consumption because a significant increase in biomass was still detected at 75 mg N ([26]). These conflicting results for Pinus spp. suggest that a wider range of nutrient availability should be tested to more accurately quantify N luxury consumption under varying cultural systems.
In recent years, there has been a trend to determine nursery fertilization in light of field performance on a specific site condition as well as seedling biomass and nutrient reserves accumulated during nursery growth ([12], [13], [27], [19], [47]). For example, in very dry Mediterranean environments some studies showed that seedlings with low nutrient concentration or content performed better ([42], [43], [7]), whereas other studies indicated that increased tissue nutrient reserves improves seedling field performance ([22], [8], [48], [29]). These contrasting results suggest that insight into nursery fertilization and field outplanting should be further tested, in particular with relation to climatic planting conditions or soil characteristics. In the present study, greater plant N reserves induced by the highest exponential fertilization rate did not promote survival and growth performance in the following two growing seasons. In contrast, the lowest pre-hardening fertilization rate (40 mg) yielded desirable growth performance. These findings suggest that, under the relatively dry conditions of our study, rational reduction of nursery fertilization might be a choice to improve seedling outplanted performance once these nursery fertilization rates are within the luxury range.
Effect of fall fertilization on nursery and field performance
The benefit of fall fertilizer on enhancement of foliage N concentration was found in Chinese pine as well as other tree seedlings ([23], [39], [16], [13], [19]). This result is consistent with the goal of fall fertilization to nutrient load by enhancing foliar N concentration ([9]). Increased foliar N concentration in fall-fertilized Chinese pine seedlings did not facilitate foliar N content, in accordance with that reported for holm oak seedling ([28]) but in contrast to results for Douglas-fir, loblolly pine, and Norway spruce seedlings ([23], [39], [13]). Lack of increased foliar N content could be attributed to the absence of increasing foliar biomass of fall-fertilized seedlings, as per other previous studies ([16], [28]).
The present and earlier findings of fall fertilization indicated mixed survival and growth responses to increased foliar N concentration. Fall fertilization had no effect on survival of Chinese pine, corresponding to results for Norway spruce, Douglas-fir, and Lutz spruce ([46], [13], [19]). However, reduced survival was reported in fall-fertilized longleaf pine (Pinus palustris Mill.) seedlings ([38]). Pre-planting diameter and height of fall-fertilized seedlings was similar to that of non fall-fertilized seedlings for Chinese pine and several other species ([24], [16], [13], [28]). These trends were maintained in first-year diameter for Chinese pine, which was contrary to that reported for slash pine seedlings ([16]). For height, favorable effects of fall fertilization did not appear until the second growing season in our study, despite frequent reports of improved first-year height growth in fall fertilized seedlings ([24], [16], [13]). Inconclusive growth performance across these studies again demonstrates that comprehensive consideration of both nursery and field performance determines rational N supply in the nursery for seedling production to meet a particular site condition. Overall, 12 and 24 mg N fall fertilization rates are more effective during hardening for Chinese pine seedling in terms of foliar N concentration prior to planting, and total height and height increment at the second year after outplanting.
Interaction between pre-hardening and fall fertilization on nursery seedling N storage and subsequent field performance
In previous studies investigating fall fertilization, seedlings generally received the same dose of pre-hardening fertilization and were subsequently fall-fertilized at multiple treatment rates ([46], [16], [13], [19]). This precludes the ability to distinguish how pre-hardening fertilization affects fall fertilization; and the use of a single pre-hardening fertilization rate potentially contributes to the inconsistent results of fall fertilization among studies. For black spruce seedlings, interactions influenced both N concentration and content of the whole plant ([4]). Interaction only occurred in root N concentration of Chinese cork oak seedlings ([21]) and in N content of the whole plant for Chinese pine in our study. Additionally, fall fertilization had adverse effects on N content when seedlings were fertilized at the pre-hardening rate of 120 mg N, indicating that this rate may have the approximated optimal nutrient input whereas additional fertilization has proven to induce some toxicity ([18]).
First-year height after outplanting indicated that fall fertilization had a slightly advantageous effect under 40 and 80 mg N per seedling of pre-hardening fertilization regimes, whereas negative impacts were observed at the highest rate. This could be related to incipient toxicity for seedlings fertilized at 120 mg that were subsequently fertilized with additional N in the fall. This interactive effect of pre-hardening and fall fertilization was transitory and vanished at the second-year of outplanting.
Conclusions
Based on nursery seedling growth and N content during the pre-hardening period, the three pre-hardening fertilization rates (40, 80 and 120 mg N) induced luxury consumption of Chinese pine seedlings. Fall fertilization of seedlings that received 40-80 mg N per seedling during pre-hardening further N loaded Chinese pine. For the highest pre-hardening fertilization rate, however, fall fertilization decreased N content.
Maximum diameter was observed in the lowest nursery exponential fertilization rate at the second year after outplanting. Supplemental fertilization during fall with 12 and 24 mg N per seedling were more effective in enhancing both total height and height increment at the second year after outplanting. Considering nursery and field performance, fertilization cost and potential nitrate leaching risks, combining pre-hardening fertilization with fall fertilization may serve as a more effective means to promote N storage and outplanting growth as compared with pre-hardening fertilization alone. Further research is needed, however, to optimize application rates under this fertilization scheme for Chinese pine and other important reforestation species under varying cultural regimes.
Acknowledgements
The study was funded by the Fundamental Research Funds for the Central Universities, China (contract no. TD2011-08). We specially thank the Meteorological Bureau of Pingquan County for assistance with providing weather data.
References
Gscholar
Gscholar
CrossRef | Gscholar
Online | Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Jiaxi Wang
Key Laboratory for Silviculture and Conservation, Ministry of Education, Beijing Forestry University, 35 East Qinghua Road, Haidian District, Beijing 100083 (China)
Department of Natural Systems and Resources, Universidad Politécnica de Madrid, Ciudad Universitaria s/n, 28040 Madrid (Spain)
Department of Forestry and Natural Resources, Hardwood Tree Improvement and Regeneration Center, Purdue University, West Lafayette, IN 47907-2061 (USA)
Corresponding author
Paper Info
Citation
Li G, Wang J, Oliet JA, Jacobs DF (2016). Combined pre-hardening and fall fertilization facilitates N storage and field performance of Pinus tabulaeformis seedlings. iForest 9: 483-489. - doi: 10.3832/ifor1708-008
Academic Editor
Gianfranco Minotta
Paper history
Received: May 12, 2015
Accepted: Sep 20, 2015
First online: Jan 07, 2016
Publication Date: Jun 01, 2016
Publication Time: 3.63 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2016
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 40374
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 34243
Abstract Page Views: 2094
PDF Downloads: 3048
Citation/Reference Downloads: 19
XML Downloads: 970
Web Metrics
Days since publication: 3032
Overall contacts: 40374
Avg. contacts per week: 93.21
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Feb 2023)
Total number of cites (since 2016): 10
Average cites per year: 1.25
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Combined effects of short-day treatment and fall fertilization on growth, nutrient status, and spring bud break of Pinus tabulaeformis seedlings
vol. 10, pp. 242-249 (online: 11 February 2017)
Technical Reports
Nursery practices increase seedling performance on nutrient-poor soils in Swietenia humilis
vol. 8, pp. 552-557 (online: 09 December 2014)
Research Articles
Substrates and nutrient addition rates affect morphology and physiology of Pinus leiophylla seedlings in the nursery stage
vol. 10, pp. 115-120 (online: 02 October 2016)
Research Articles
Nursery fertilization affected field performance and nutrient resorption of Populus tomentosa Carr. ploidy levels
vol. 15, pp. 16-23 (online: 24 January 2022)
Research Articles
Conservation of Betula oycoviensis, an endangered rare taxon, using vegetative propagation methods
vol. 13, pp. 107-113 (online: 23 March 2020)
Research Articles
Linking nursery nutritional status and water availability post-planting under intense summer drought: the case of a South American Mediterranean tree species
vol. 9, pp. 758-765 (online: 03 June 2016)
Research Articles
Forecasting the field performance of Austrian pine seedlings using morphological attributes
vol. 10, pp. 99-107 (online: 13 October 2016)
Research Articles
Controlled-release fertilizers combined with Pseudomonas fluorescens rhizobacteria inoculum improve growth in Pinus halepensis seedlings
vol. 8, pp. 12-18 (online: 12 May 2014)
Research Articles
The effect of calcium on the growth of native species in a tropical forest hotspot
vol. 11, pp. 221-226 (online: 01 March 2018)
Research Articles
Wood production and nutritional status of Pinus taeda L. in response to fertilization and liming: a meta-analysis of the Americas
vol. 16, pp. 195-201 (online: 25 July 2023)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







