
Influence of mother plant and scarification agents on seed germination rate and vigor in Retama sphaerocarpa L. (Boissier)
iForest - Biogeosciences and Forestry, Volume 7, Issue 5, Pages 306-312 (2014)
doi: https://doi.org/10.3832/ifor1155-007
Published: Apr 08, 2014 - Copyright © 2014 SISEF
Research Articles
Abstract
The aim of this study was to optimize nursery production of Retama sphaerocarpa, an important shrub species in some plant communities of arid and semi-arid zones, which is used in the restoration of degraded areas. This Fabaceae family species has seed coat dormancy, which may hinder germination. In order to assess the influence of scarification agents and mother plant effect on germination rate and seedling vigour, a germination test with fresh seed was performed in a germination chamber, following a factorial design with two fixed factors (mother plant effect and scarification agent). The test took place in the forest nursery of the Institute of Agricultural Sciences, Lisbon, Portugal. The fresh seeds were subjected to scarification by two agents, sulphuric acid (96% concentration) and hot water (80°C), the former for 60 minutes and the latter for 30 seconds, plus an untreated control. The scarification of seeds using sulphuric acid was found to be the best method to enhance germination of this species (p<0.05), as it allowed to reach the highest germination rate (>79%) and the best vigour index. Differences between mother plants were also statistically significant and could influence germination success. We suggest that effectiveness of hot water scarification may be improved by additional time of seed immersion. The technical viability for seed production of this species was confirmed and key factors for its success identified. Owing to its potential for use on land rehabilitation, increment on plant commercial demand should be expected in the near future.
Keywords
Fresh Seed, Germination, Mother Plant, Retama sphaerocarpa, Scarification, Vigor
Introduction
Arid and semi-arid zones are very susceptible to anthropogenic disturbance and degradation ([19], [39]). Usually plant cover is scarce in these areas, being susceptible to disturbance and prolonged drought periods ([39]). Also, human activities often impoverish natural seed banks, negatively influencing the regeneration capacity of arid and semi-arid ecosystems ([5], [8]). Therefore, natural regeneration of Mediterranean woodlands after disturbance is often absent or very slow ([38], [39]). Consequently, the area occupied by such ecosystems has decreased considerably over the centuries, particularly in the driest areas (e.g. [36], [38], [4], [30]). The effects of non-sustainable land management practices, such as intensification of grazing, could limit successional patterns and vegetation responses to natural disturbance ([16]). Nitrogen cycling may be a key factor in those processes, as well as the occurrence of ruderal plants with phytotoxic capabilities that inhibit other plant species. The establishment of a nitrogen-fixing plant prone to play a facilitation role in disturbed woodlands, like Retama sphaerocarpa (L.) Boissier, may represent an opportunity to rehabilitate degraded communities ([31]).
Suitable management and restoration activities in Mediterranean woodlands are fundamental, in that vegetation cover may mitigate erosion and desertification effects, preserving soil quality and favoring soil evolution ([7], [26]). Such activities imply the diversification of plant species used in restoration projects, a broad use of the wide variety of native species available, and the consideration of wildlife, microorganisms, soil and flora as an integrated framework ([39]).
Retama sphaerocarpa (Fabaceae family) is a shrub naturally growing in the above mentioned ecosystems of northern Africa and the Iberian peninsula. It is a common species in Portugal, mainly in the southern part of the country, although its natural distribution includes much of central Portugal and some areas in the interior north deep valleys ([3]). This species is well adapted to the seasonal water stress conditions and shallow soils typical of semi-arid Mediterranean ecosystems, showing morphological and architectural features suitable to water conservation, such as small leaf area (very small leafs, only present for a short period during the spring) and the presence of photosynthetic stems with a vertical orientation that minimize light interception and overheating ([12]). Additionally, it has a thick root system, which can reach 25 meters deep through crevices in the soil parent material ([12]). It also has the ability to hydraulically lift water from deep to shallow soil layers ([27]).
R. sphaerocarpa exhibits a broad ecological range with no specific soil type preference, although it suffers much hardship in very cold or humid locations. It can be found in the Iberian Peninsula from sea level up to 1400 meters of elevation ([33], [37], [17]).
As many other woody species typical of the Mediterranean region and other semi-arid zones, Retama sphaerocarpa grows for a short period of time in the spring, with favorable temperature and soil moisture conditions ([13]). Fruit production is often abundant, normally occurring between the end of July and the beginning of August. The seeds have a hard seed coat, measuring approximately 3.5-5 x 5-8 mm ([37]) and weighting between 90 and 100 mg ([29]). The main seed dispersal agent is the wind, although water and sometimes animals are also involved in the dispersal process ([13], [20], [29]).
Retama sphaerocarpa plays an important role in the restoration of degraded areas, due to its capability to withstand dry environments, the symbiosis between its root system and nitrogen-fixing bacteria, its phytoremediation capabilities, and also its importance in some plant communities of arid and semi-arid zones ([2], [6], [22]). Moreover, it has some potential as a cultivated crop for energy production, for carbon sequestration and for ornamental purposes in Mediterranean gardens ([33], [1], [34]).
Similar to many Mediterranean shrub species ([26]), information on Retama sphaerocarpa propagation in the literature is scarce and often inaccurate, likely due to the physical dormancy of its seeds that hinders the germination process ([29]). Under natural conditions dormancy allows the persistence of viable seeds within the soil seed bank over long time periods ([40]). Seed hardiness gradually declines over time as a result of natural factors, which may include the friction with soil particles (mechanical scarification), wildfire heat (thermal scarification), or the action of digestive acids when seeds are eaten by wildlife and/or domestic animals (chemical scarification - [40], [14], [32], [11]). The persistence on the soil seed bank allows the survival of some viable seeds until the occurrence of environmental conditions favorable to germination, plant survival and growth ([40], [18], [28]).
Several pre-germination treatments may be used in the nursery to emulate the mentioned natural processes, such as the immersion in near-boiling water or sulphuric acid, the physical abrasion or pressure, or the seed exposure to low or alternating temperatures ([14]). López et al. ([18]) reports germination rates after 30 days of 40-47% (after a pre-germination treatment of 1 minute in 100 °C water, simulating natural warming due to wildfires), 97-100% (following 60 minutes in sulphuric acid, 96% concentration, simulating animal consumption) and 0% (untreated seed). These values refer to seed collected in the same year and stored in paper bags at room temperature. On the other hand, Ruiz de la Torre et al. ([33]) indicates germination rates of 75-85% (30 seconds in 80 °C water, followed by cooling in cold water) and of 35% (untreated seed), not mentioning the duration of the germination tests. Pérez-Fernández & Rodríguez-Echeverría ([23]) report germination percentages after 35 days of 5% (seeds pre-soaked in boiling water, followed by cooling for 8 hours) and 20% (after similar pre-soaking followed by 10 min of smoke exposure to simulate the chemical action of wildfire smoke).
The aim of this study was to investigate the germination performance of Retama sphaerocarpa seeds from a Portuguese provenance, with the objective of improving nursery production techniques and better understanding its seed germination characteristics, as well as its suitability for the ecological restoration of Mediterranean ecosystems and its exploitability as a bioenergy crop.
Materials and methods
Seed collection and handling
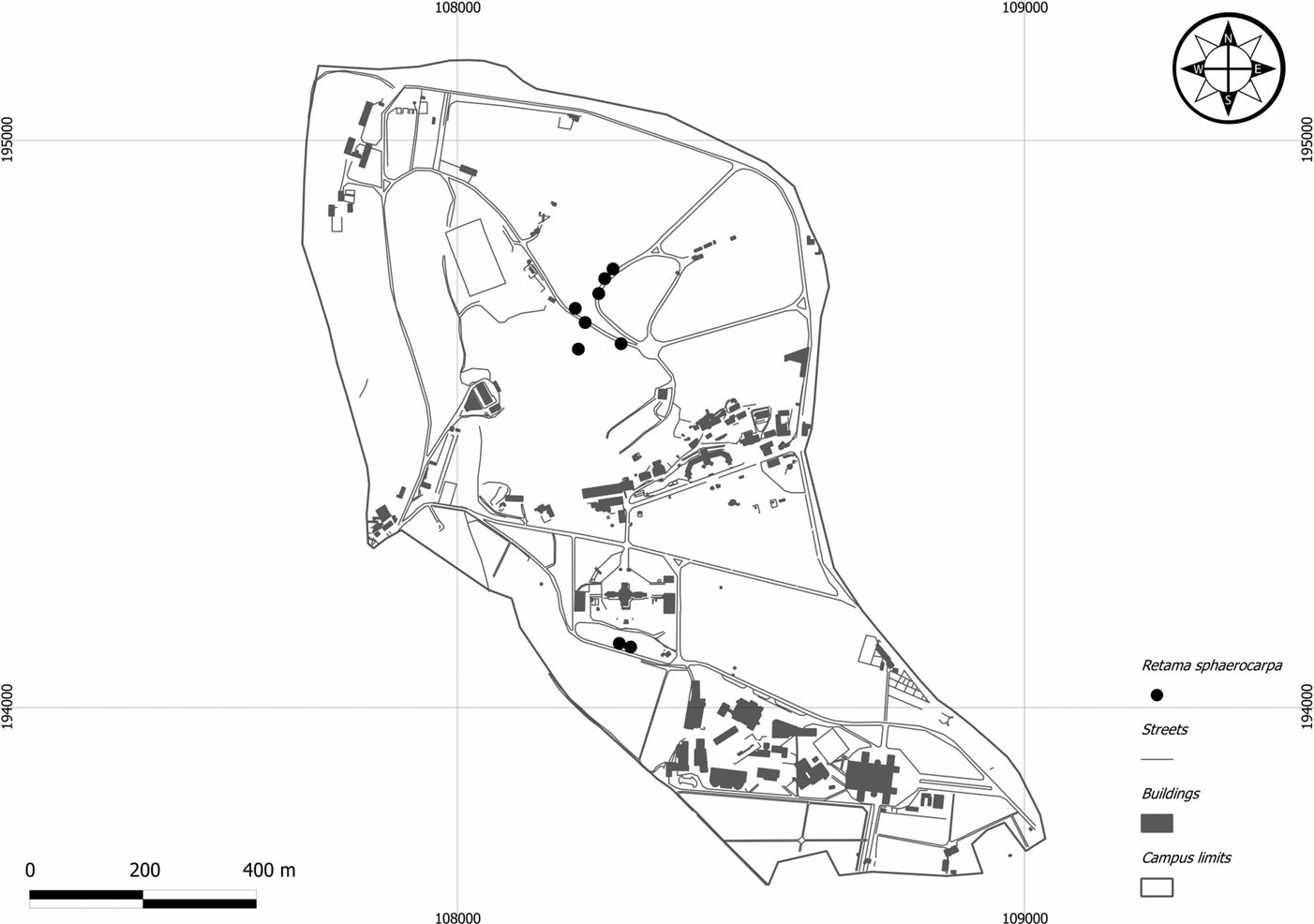
Seeds were hand-harvested at the end of September 2010 from 9 different mother plants at least 20 meters apart located in the 100-ha university campus (Fig. 1) of the Institute of Agricultural Sciences at Tapada da Ajuda (Lisbon, Portugal - 38°42’27.61” N, 9°10’56.32” W, max elevation: 131 m a.s.l.). The sampled mother plants belong to a natural population located in Tapada da Ajuda and in the adjacent Monsanto peri-urban county forest.
Fig. 1 - Map of the Tapada da Ajuda University Campus (Lisbon, Portugal). Black dots represent the mother plants of Retama sphaerocarpa sampled for seed germination analysis.
Collected seeds were kept in their pods to avoid external contamination and stored in kraft paper envelopes at room temperature until the start of the trial. All damaged or unripen seeds were rejected upon shelling.
Experimental design
The germination trials started in the beginning of October 2010. Seeds were disinfected by immersion in a 1% sodium hypochlorite solution for 10 minutes and thoroughly rinsed with distilled water ([32]).
A factorial design with two fixed factors (mother plant effect and scarification agent) was used. Nine levels were considered for mother plant effect, and the following three levels for the scarification procedure: (A) control with no scarification agent; (B) scarification by soaking the seeds in a water solution containing 96% sulphuric acid for 60 minutes in the dark ([18]); (C) scarification by soaking the seeds in hot water (80 °C) for 30 seconds, followed by cooling down at room temperature in distilled water ([33]). A total of 27 different treatments (9 mother trees x 3 scarification procedures) were considered. Each treatment had 3 replications per mother plant (3 samples with 20 seeds each), totaling 60 seeds per each mother plant and treatment. Overall, a total of 1620 seeds were used.
No specific ISTA (International Seed Testing Association) rules do exist for Retama sphaerocarpa, so ISTA rules for Cytisus spp. were used as for substrate, temperature and trial duration ([15]). However, ISTA recommendations on the mechanical scarification of seeds for breaking dormancy in the latter species were not applied, due to the small size of Retama sphaerocarpa seeds and the time consumption of their scarification without suitable devices.
The germination test was carried out in modified Jacobsen’s individual apparatus trays ([40]) placed into a germination chamber at a temperature of 21 ± 3 °C, with no moisture control and in the dark. Seed watering was carried out whenever needed (every 2-3 days, on average) by placing the seeds on a filter paper moistened with distilled water.
Data analysis
Seeds showing an emerging radicle ≥ 1 mm were considered as germinated, then counted and removed from the trays ([18]). Seeds showing fungal contamination on the whole coat surface were removed and considered as non-germinated.
The number of germinated seeds for each treatment was recorded, and the corresponding germination percentages obtained. Germination vigor index (V) was calculated for each treatment as a measure of the germination rate per unit time, following the method proposed by López et al. ([18]):
where a, b, c, … represents the number of germinated seeds after 1, 2, 3, … days, respectively; x is the number of seeds germinated at the end of the trial (n-th day); and S is the total number of seeds in the survey. The index values range between 0 and 100 (null to very fast), and were categorized as reported in Tab. 1 ([18]).
Tab. 1 - Classes of the germination vigor index (V), as proposed by López et al. ([18]).
| Category | Value range |
|---|---|
| Very fast | 33.33 ≤ V ≤ 100 |
| Fast | 11.11 ≤ V <33.33 |
| Average | 5.0 ≤ V <11.11 |
| Slow | 0.0 < V <5.0 |
| Null | V = 0.0 |
Statistical analysis
All statistical analyses were carried out by the use of the SPSS® software package (PASW Statistics, v. 18.0.0, 2009).
Departure of the germination data from normal distribution was tested by applying the D’Agostino-Pearson’s test ([9]), using a SPSS-macro developed by DeCarlo ([10]). Given the assumptions for the analysis of variance were not met after data transformation, suitable non-parametric tests were used.
To ascertain for differences in germination and germination vigor index among scarification procedures, a Kruskal-Wallis test was carried out. Significance of pairwise differences between treatments was assessed by Games-Howell’s multiple comparisons test using mean ranks ([35]). Finally, Mann-Whitney U-test was used to assess the statistical significance of differences in the germination and germination vigor index among mother plants.
Results
Scarification effect on germination and vigor
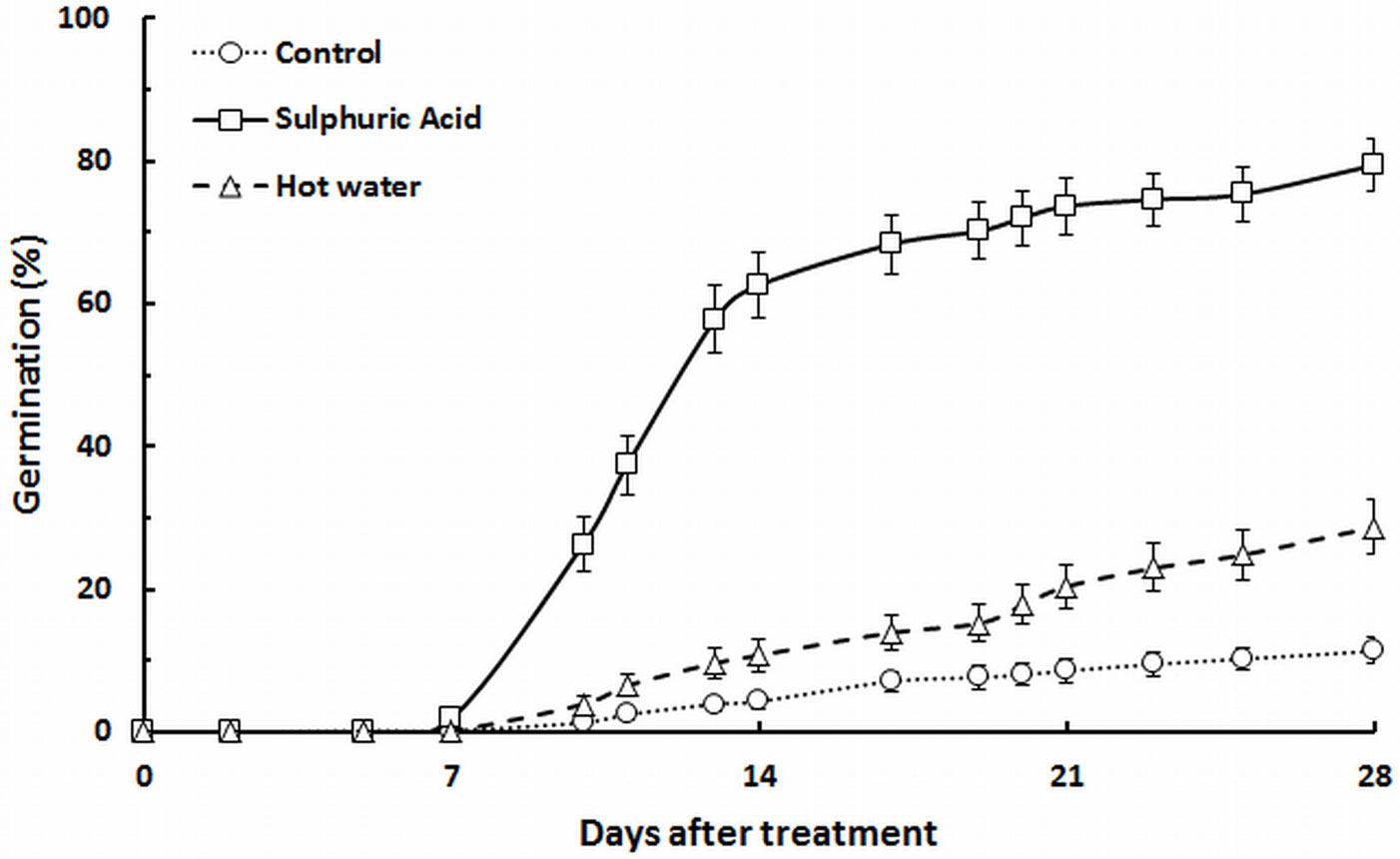
After 28 days of survey, seeds scarified with sulphuric acid (treatment B) showed the best results, with a germination percentage of 79.4 ± 3.8 % (mean ± standard error over the 9 mother plants). Germination success for seeds scarified with hot water (treatment C) was only 28.7 ± 3.8 %. Control treatment showed the lowest germination percentage (only 11.3 ± 1.8 %). According to vigor index categories reported in Tab. 1, germination was classified as “very fast” for treatment B, “average” for treatment C and “slow” for treatment A. Tab. 2 displays the mean values of germination percentages and germination vigor index obtained for each treatment. The cumulative germination curves pooled over all mother plants are displayed for each treatment in Fig. 2.
Tab. 2 - Mean ± standard error of germination percentages and germination vigor index for seeds subjected to three treatments after 28 days of survey. Different letters indicate significant pairwise differences between treatments after Games-Howell’s test (p<0.05).
| Treatment | Number of seeds | Germination percentage |
Germination vigor index |
|---|---|---|---|
| (A) Control | 540 | 11.3 ± 1.8 a | 3.85 ± 0.75 a |
| (B) Sulphuric Acid | 540 | 79.4 ± 3.8 b | 39.13 ± 2.47 b |
| (C) Hot water | 540 | 28.7 ± 3.8 c | 9.00 ± 1.41 c |
Fig. 2 - Cumulative distribution of the germination percentage in each treatment for Retama sphaerocarpa seeds. Vertical bars are standard errors (n=27).
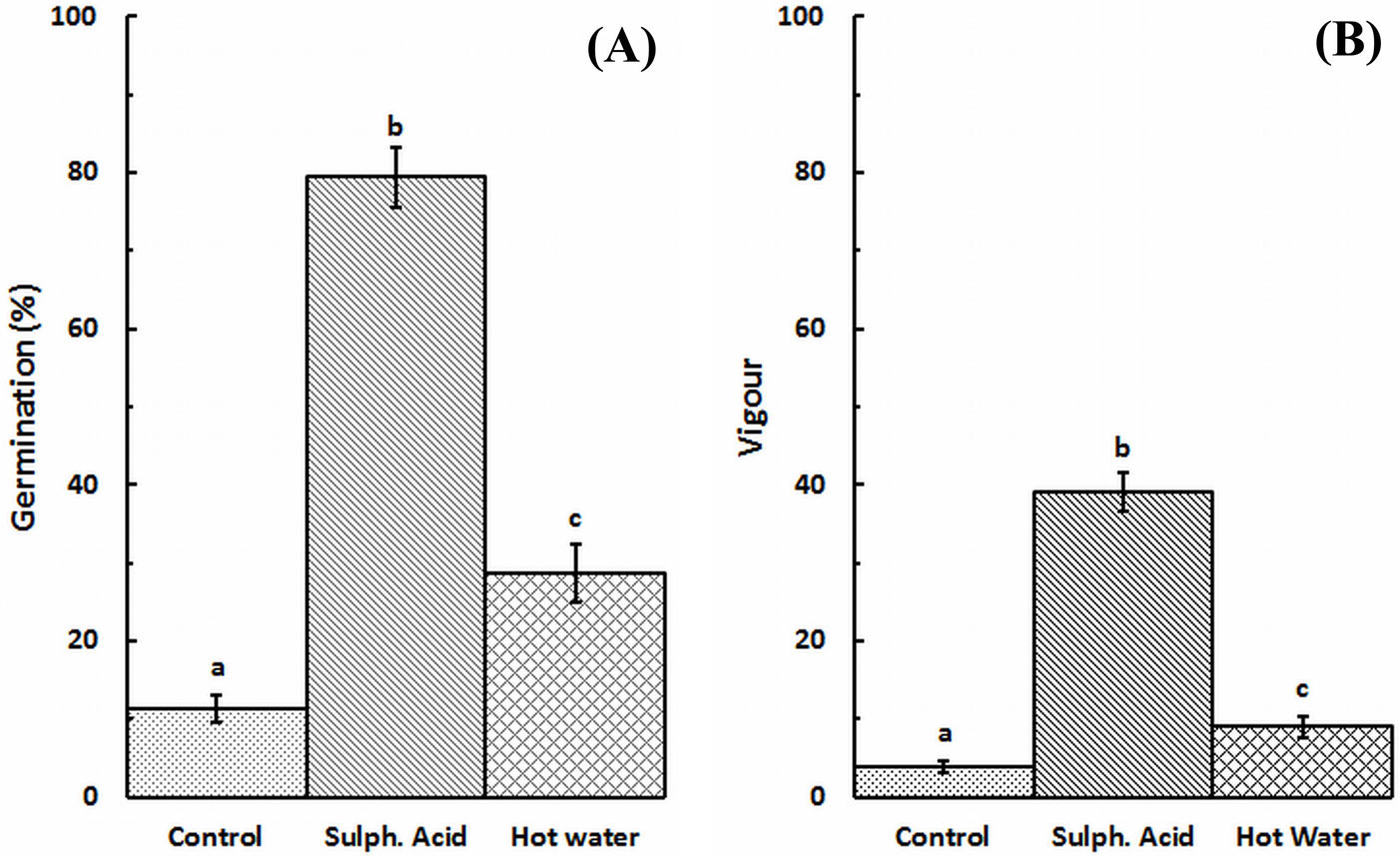
Both germination rate and germination vigor index were significantly different among treatments after Kruskal-Wallis test (H[2] = 52.494, p = 0.000, n = 81; and H[2] = 52.532, p = 0.000, n = 81; respectively). Moreover, significant pairwise differences were found between all treatments after Games-Howell’s test (p≤0.05 - Fig. 3).
Fig. 3 - Germination rate (A) and germination vigor index (B) by treatment. Vertical bars are standard errors (n=27). Different letters indicate significant pairwise differences between treatments after Games-Howell’s test (p<0.05).
Mother plant effect on germination and vigor
Germination percentages were broadly similar among seed lots from all mother plants, with the exception of mother plant I1 and, to a lesser extent, mother plants I8 and I9. The highest germination percentage pooled over the 3 treatments was obtained for the seed lot from mother plant I7 (52.2 ± 9.8 %) and the lowest from mother plant I1 (18.3 ± 6.0%). According to vigor index categories reported in Tab. 1, germination was classified as “fast” for all seed lots, except for those from mother plant I1 which was considered as “average”. Tab. 3 displays the germination percentage and germination vigor index of the different mother plants. The cumulative germination curves pooled over all treatments are displayed by mother plant in Fig. 4.
Tab. 3 - Mean ± standard error of germination percentages and germination vigor index for seeds collected from 9 mother plants after 28 days of survey. Different letters indicate significant pairwise differences between mother plants after Mann-Whitney U-test (p<0.05).
| Mother plant |
Number of seeds |
Germination percentage |
Germination vigor index |
|---|---|---|---|
| I1 | 180 | 18.3 ± 6.0a | 5.95 ± 2.22a |
| I2 | 180 | 48.4 ± 9.6b | 24.50 ± 5.91b |
| I3 | 180 | 47.2 ± 9.6b | 20.09 ± 5.06b |
| I4 | 180 | 40.6 ± 12ab | 17.37 ± 6.09ab |
| I5 | 180 | 42.9 ± 9.8ab | 20.05 ± 6.28ab |
| I6 | 180 | 40.0 ± 11.6ab | 16.82 ± 5.80ab |
| I7 | 180 | 52.2 ± 9.8b | 17.53 ± 5.20b |
| I8 | 180 | 34.4 ± 15.2ab | 17.26 ± 8.31ab |
| I9 | 180 | 34.4 ± 15.2ab | 16.40 ± 7.76ab |
Fig. 4 - Germination curves of Retama sphaerocarpa by seed lots from different mother plants. Vertical bars are standard errors (n=3).
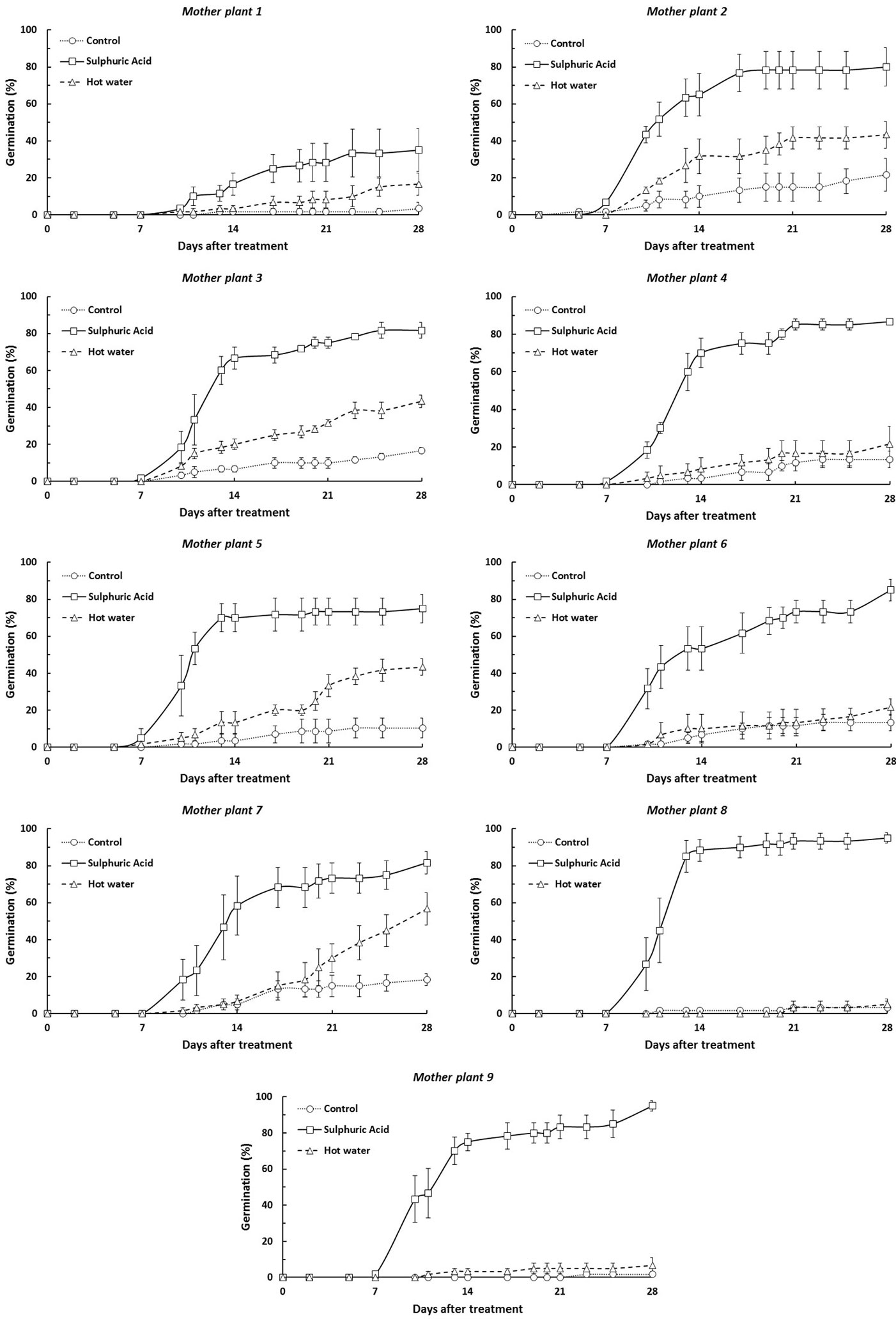
Significant differences in germination rate were found between three pairs of mother plants after Mann-Whitney U-test: I1 and I2 (U=13, p<0.05), I1 and I3 (U=15.5, p<0.05) and between I1 and I7 (U=14, p<0.05 - Fig. 5).
Fig. 5 - Germination rate (A) and germination vigor index (B) by mother plant. Vertical bars are standard errors (n=9). Different letters indicate significant pairwise differences between mother plants after Mann-Whitney U-test (p<0.05).
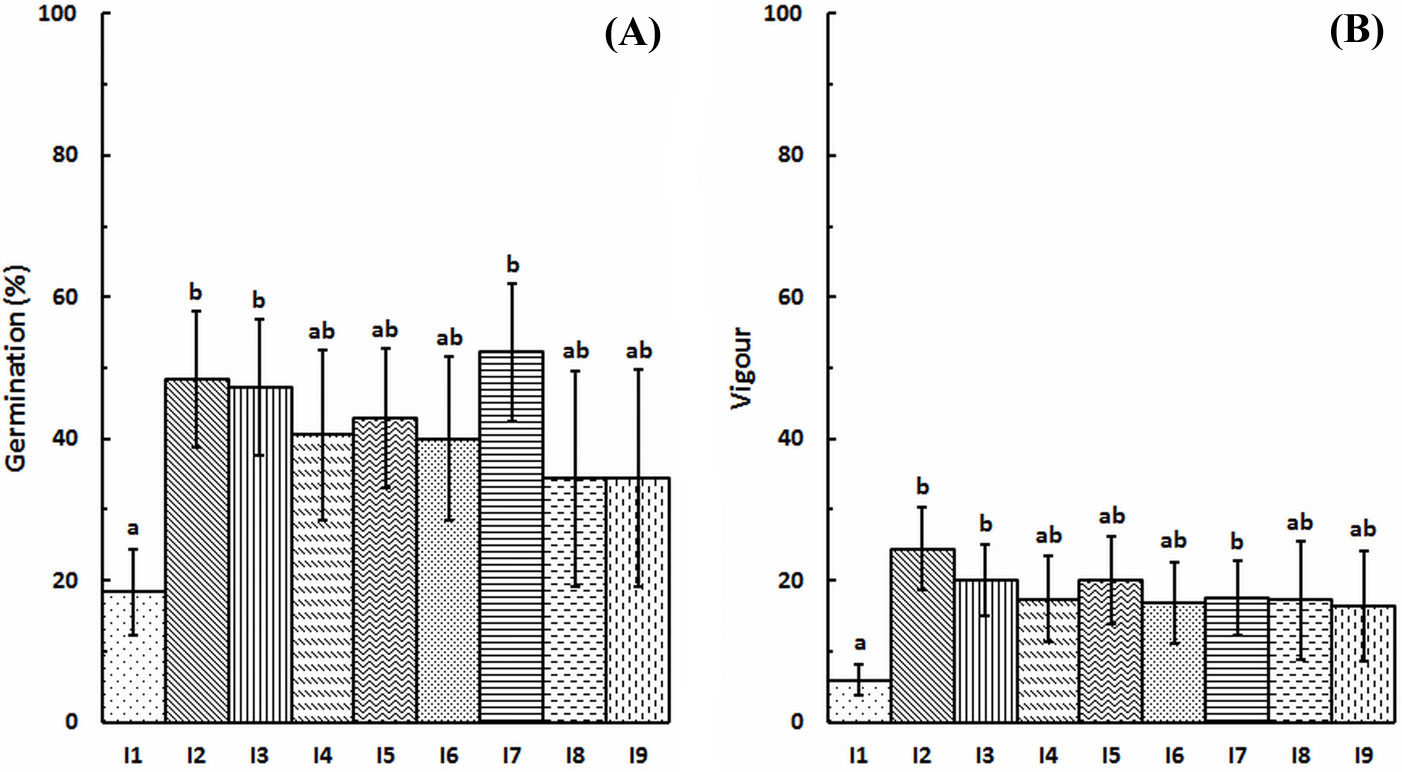
Germination vigor index was significantly different only between two pairs of mother trees: I1 and I2 (U=12.5, p<0.05), I1 and I3 (U=16, p<0.05 - Fig. 5).
Discussion
Our results clearly indicate that seed scarification is needed to improve Retama sphaerocarpa germination rate for commercial seedling production in nursery. Treatment with sulphuric acid provided the best germination results, probably due to its effectiveness in degrading the seed coat. Thermal scarification using hot water gave worse results, though in some cases its efficacy in breaking seed dormancy seems to be delayed in time (e.g., mother plant I7). As suggested by Pérez-Fernández et al. ([24]) for other Fabaceae species, stronger hot water scarification (e.g., longer seed immersion) may boost the seed germination rate also for Retama sphaerocarpa.
Similar results were obtained when the germination vigor index is considered, being the seeds treated with sulphuric acid classified in the “very fast” germination category. As reported by López et al. ([18]), seed scarification is needed not only to improve the germination capability, but also to speed up that process.
In this study, significant differences in germination rate and vigor were found among seed lots from different mother plants (I1 vs. I2, I3 and I7). Such differences may be interpreted as of genetic origin (for example, variation of seed coat thickness among seed lots from different mother plants), or may be related with the vitality of the mother plants which may influence seed maturation (for example, different exposure to light of mother plants related with their location within the sampled area). Additional source of variation in seed germination rate may be related to the age of the collected pods. In fact, Retama shpaerocarpa shrubs tend to keep some seed pods for more than one year ([13]), making such seeds more prone to fungal contamination, thus reducing their germination capacity. Indeed, some fungal contamination of seeds was observed all over the germination survey carried out. The most susceptible treatment to fungal infection was the thermal scarification with hot water (data not shown), but also seed lots from single mother plants (namely 1 and 5) were clearly more affected than others by contamination. Such susceptibility to infection could have possibly generated further variation in our dataset, so results (especially differences in germination among seed lots from different mother plants) should be interpreted with caution.
Seed coat thickness in legumes may considerably vary ([25]). As a consequence, the treatments applied in this study might provoke an unwanted selection, destroying seeds with lighter coats and simultaneously not scarifying seeds with thicker coats ([25]). It has been reported mechanical scarification as a safer and less damaging alternative to chemical or thermal treatments ([25]). Moreover, the use of percussion to break seed physical dormancy has been proposed as a further alternative, as successfully tested on several herbaceous Fabaceae by Mondoni et al. ([21]). However, mechanical scarification of large amounts of seed (as in commercial seedling production in nursery) requires specific, highly expensive machinery. On the other hand, handling of sulphuric acid or large amounts of boiling water requires special safety measures and skilled workers.
Our results are slightly different from those reported in the literature: only the treatment with sulphuric acid obtained germination percentages in line with expectations (79.4 ± 3.8 %). However, several authors do not mention the duration of their experiments or the length of seed storage (e.g., [33], [23]), making difficult the comparison. Several reasons could be invoked to explain the differences mentioned above: (i) the presence in the collected material of seeds at different stages of maturity; (ii) differences in the germination capacity among seed lots from different mother plants; (iii) differences in seed morphology (seed coat thickness); and (iv) differences in fungal contamination of the seed lots.
Retama sphaerocarpa has a great potential for use in restoration of degraded ecosystems and production of biomass for energy purposes, as well as for low-maintenance Mediterranean gardens. Thus, future demand for this species is expected to increase, justifying its production by commercial plant nurseries.
Conclusions
Seed scarification using 96% sulphuric acid for 60 minutes in the dark has proven the most effective pre-germination treatment for breaking dormancy of Retama sphaerocarpa seeds, enabling the maximization of the number of germinated seeds and ensuring shorter germination times. This allows a greater age uniformity of the plant batch, which eases the management and disposal of the nursery stocks.
Pre-germination treatments with hot water may provide similar results in the future, but additional tests are recommended using longer soaking times and/or keeping the seeds immersed while the water cools down, with the main advantage of cheaper costs and safer procedures in the nursery.
Mechanical scarification might be a safer and less damaging alternative to the above treatments, although requiring suitable (and costly) machinery for handling large quantities of seed.
Genetic or vigor-related characteristics of the mother plants of Retama sphaerocarpa may influence the effectiveness of pre-germination treatments. Finally, more information is needed on the germination capacity of stored seed, and on different seed storage methods for this species.
Acknowledgments
This study has been funded by the Forest Research Center (Pest-OE/AGR/UI0239/ 2011). Technical support was provided by the project “Enerwood - Potential of native woody species for energy production from biomass” (PTDC/AGR-CFL/114826/2009). The authors would like to thank Regina Fabião for assisting in seed harvesting and Rita Moreira and Marta Carneiro for their help in seed shelling. We also thank an anonymous referee for helpful comments on the manuscript.
References
Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Carla Faria
Maria Helena Almeida
António Fabião
Centro de Estudos Florestais, Instituto Superior de Agronomia, Universidade de Lisboa, Tapada da Ajuda, P-1349-017 Lisboa (Portugal)
Corresponding author
Paper Info
Citation
Fabião A, Faria C, Almeida MH, Fabião A (2014). Influence of mother plant and scarification agents on seed germination rate and vigor in Retama sphaerocarpa L. (Boissier). iForest 7: 306-312. - doi: 10.3832/ifor1155-007
Academic Editor
Gianfranco Minotta
Paper history
Received: Oct 18, 2013
Accepted: Jan 20, 2014
First online: Apr 08, 2014
Publication Date: Oct 01, 2014
Publication Time: 2.60 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2014
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 59365
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 49017
Abstract Page Views: 3981
PDF Downloads: 4747
Citation/Reference Downloads: 41
XML Downloads: 1579
Web Metrics
Days since publication: 4422
Overall contacts: 59365
Avg. contacts per week: 93.97
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2014): 3
Average cites per year: 0.25
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Conservation of Betula oycoviensis, an endangered rare taxon, using vegetative propagation methods
vol. 13, pp. 107-113 (online: 23 March 2020)
Research Articles
Role of serotiny on Pinus pinaster Aiton germination and its relation to mother plant age and fire severity
vol. 12, pp. 491-497 (online: 02 November 2019)
Research Articles
Effects of brassinosteroid application on seed germination of Norway spruce, Scots pine, Douglas fir and English oak
vol. 10, pp. 121-127 (online: 02 October 2016)
Technical Reports
Effects of different mechanical treatments on Quercus variabilis, Q. wutaishanica and Q. robur acorn germination
vol. 8, pp. 728-734 (online: 05 May 2015)
Research Articles
The effectiveness of short-term microwave irradiation on the process of seed extraction from Scots pine cones (Pinus sylvestris L.)
vol. 13, pp. 73-79 (online: 13 February 2020)
Research Articles
Germination and seedling growth of holm oak (Quercus ilex L.): effects of provenance, temperature, and radicle pruning
vol. 7, pp. 103-109 (online: 18 December 2013)
Research Articles
Can we expect more from Alnus glutinosa (L.) Gaertn. in a changing climate?
vol. 19, pp. 69-76 (online: 08 March 2026)
Research Articles
Effects of different nut pretreatments and substrates on germination and seedlings growth of Neocarya macrophylla Sabine in Basse Casamance, Senegal
vol. 17, pp. 346-352 (online: 03 November 2024)
Short Communications
Evidence of Alectoris chukar (Aves, Galliformes) as seed dispersal and germinating agent for Pistacia khinjuk in Balochistan, Pakistan
vol. 14, pp. 378-382 (online: 22 August 2021)
Research Articles
The maternal environment of European beech (Fagus sylvatica L.) affects intrapopulation variability in seed traits and germination
vol. 19, pp. 114-121 (online: 10 April 2026)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







