
Growth dynamics and productivity of pure and mixed Castanea sativa Mill. and Pseudotsuga menziesii (Mirb.) Franco plantations in northern Portugal
iForest - Biogeosciences and Forestry, Volume 7, Issue 2, Pages 92-102 (2014)
doi: https://doi.org/10.3832/ifor1087-007
Published: Dec 18, 2013 - Copyright © 2014 SISEF
Research Articles
Abstract
Since the late 1980’s the productivity of monocultures versus mixed-species forests has been the object of special interest and study by forest managers and ecologists. All over the world mixed plantations have been established in different proportions to analyse if mixtures can provide greater yields and more benefits than monocultures of the component species and also to understand if they can be an interesting economic option. An experimental design trial was set up in the north of Portugal in a replacement series with pure and mixed Castanea sativa Mill. and Pseudotsuga menziesii (Mirb.) Franco. The objective of this study was to assess growth dynamics and compare the aboveground biomass and net primary production of the two species in pure and mixed treatments in proportions 1:1 and 1:3. The growth was measured at 7, 11, 15, 17, 19, 27 and 28 years after planting and aboveground net primary production was estimated at age 28 years. As a component of the mixed treatments, P. menziesii exhibited greater height, diameter and aboveground biomass than C. sativa. Relative yield total indicated a higher productivity in the mixtures compared with the pure treatments. Early in the development the pure treatments had a similar aboveground biomass per hectare as the mixtures, but later the mixtures had more yield than the pure treatments. The mixture productivity increase trough time appears to be a result of both canopy stratification and better use of site resources. The aboveground net primary production was also higher in mixed than in the pure treatments. This study shows the importance of comparing mixed and pure stands.
Keywords
Growth, Productivity, Biomass, Mixed-species, Interactions, Replacement Series
Introduction
Mixed forest ecosystems are important to human life. Similarly to monoculture plantations, they ensure timber production but also provide other benefits through the diversification of forests products ([32], [43], [54]).
In the last few years, mixed stands dynamics returned into the focus of forest science ([18], [53]). Research in experimental mixed-species plantations has lately increased with the establishment of replicated plots of monospecific and mixed plantations at the same site, followed by extensive data collection ([3], [18], [31]). The results of this research have given a better understanding of growth dynamics, productivity and mixtures interaction over a relatively short time period.
One of the main objectives studying mixed -species plantations is to analyze whether mixed stands can provide not only greater total yields, as opposed to monocultures of equal densities, but also other benefits that may outweigh the advantages of stand management in monocultures ([3], [31]). To a large extent, superior productivity of mixed-species stands to monocultures depends on the design and proportion of the mixture, soils, silviculture, tree species and the availability of resources ([20], [8]). According to Kelty ([30]), substantial differences between species’ characteristics - in terms of shade tolerance, height growth patterns, crown structure, root depth and structure - may lead to an optimal capture and use of site resources and therefore to an overall higher productivity as compared to monocultures ([7]). Mixed stands may also experience less intense interspecific than intraspecific light competition as a consequence of differences in shade tolerance among species. Such arrangement would, in theory, maximize the use of light because of increased light interception and light-use efficiency ([30]), leading to a higher total productivity than in pure stands ([60]). This type of response has been found in studies by Kelty ([29]), Brown ([10]), DeBell et al. ([13]), Man & Lieffers ([41]) and Garber & Maguire ([21]). Forrester et al. ([18]) mentioned that canopy stratification is a key factor to ensure the coexistence of the species until the end of the rotation. Moreover, to achieve higher productivity in mixed stands, mixed species should exhibit differences in their requirements (niches) or in the way they use site resources, and/or should positively affect the growth of each other ([61]). This concept of niche separation implies that if two species are too similar in their requirements they would eventually compete intensely to exclude the other, but if competition is sufficiently weak, the two species may coexist ([27]).
The knowledge on advantages/disadvantages of mixed vs. pure stands with respect to productivity decisively influences the forest manager decision in support or against tree species poly-culture ([48]). However, sound knowledge about mixing effects even for the most common tree species combinations is rather rare and scattered. Furthermore, it is difficult to accurately predict success of mixed-species combinations and sites especially with regard to growth dynamics ([16], [18], [3], [52]).
Several studies have focused on the effects of a valuable timber species growing in mixture with a nitrogen-fixing species to investigate facilitative interactions (increased nutrient availability) between the species and resultant stand-level productivity ([6], [4], [16], [18], [34], [50]). Complementary use of resources have also been analyzed ([15], [17]). If facilitation and complementary or competitive reduction in species’ resource requirements occur simultaneously, a higher productivity is obtained by the combination of interactions among species ([15], [31], [51]). More total light interception has also been measured in mixtures ([5], [35]). Furthermore, the effect of competition between species has been analyzed to identify the key constraint ([45]). Additional improvements in mixed-species plantations compared with monocultures have been reported, such as nutrient cycling ([19]), foliar nutrients ([10], [56], [46]), soil fertility ([43]), biomass production ([7], [8], [15]) and carbon sequestration ([28], [55], [17]).
Mixed forests in northern Portugal are an important source of timber, but little is known about their productivity. Some non-native species have been used for their establishment, though information on their yield is currently lacking. It is therefore important for this region to carry out studies on the productivity of mixed-species, to evaluate stand biomass production and compare the results with monocultures of the component species, in order to guide local farmers’ decisions on their plantations.
Pseudotsuga menziesii (Mirb.) Franco is a shade-intolerant, non-native tree species introduced in Portugal in the XIX century, when it was first planted at Sintra ([23]). It has been used in plantations on mountainous areas of central and northern Portugal, showing high adaptability, fast growth, and a high potential for timber production ([36], [38]).
Castanea sativa Mill. is native to Portugal where is known since the Miocenic period and cultivated since Roman times ([37]), and has important economic, cultural and ecological functions in northern Portugal ([39]). It covers an area of 34 087 ha ([1]), mainly spread over northeastern Portugal, with the largest production area located in the Trás-os-Montes region, which represents almost 85% of the total Portuguese coverage ([22]).
Because C. sativa has a good adaptation to the shade, it can be planted in mixed-species stands as an understory level with shade-intolerant species such as P. menziesii, which tends to occupy the upper part of the canopy ([47]).
The objectives of this study were: (1) to assess differences in growth dynamics between C. sativa and P. menziesii in both pure and mixed-species plantations in a replacement series experiment; (2) to compare on site species biomass and aboveground net primary production; and (3) to estimate specific leaf area of pure and mixed plantations. Results were expected to give a better understanding of the development of these species growing in the same site, thus sharing soil and climatic conditions. The study intended also to evaluate if specific interactions intervening between C. sativa and P. menziesii would increase the overall stand productivity.
Material and methods
Experimental site
The study was carried out at an experimental site established on private land located at Bemlhevai, Bragança district (41° 24’ N, 7° 6’ W), with an elevation of 710 m a.s.l. and almost flat (2°). The ecological zone is sub-montano SA X SM, between 400 and 700 m ([2]). The soil is litholic non-humic from serecitic schist.
The climate in this region is a transition between continental and Mediterranean. The annual precipitation is 690 mm (± 8.8), of which 60% falls from October to February, and the mean annual temperature is 12.5 °C (± 1.8 - [58]).
The experiment is a replacement series design using two trees species, Castanea sativa Mill. and Pseudotsuga menziesii (Mirb.) Franco. The total stand density is constant in all the treatments but the proportions of the two component species varies. This design is often used when productivity of both species is of interest ([18]).
Experimental design
A set of twenty-one permanent plots was established in the winter of 1981 using a complete randomized block design of C. sativa and P. menziesii ([39], [37]). Each 512 m2 plot was planted using a 4 × 2 m spacing for a total of 64 trees. Plots were set up in three blocks (two species per block) with no buffer between the blocks. Each block contains seven treatments in four proportions (Fig. 1). The treatments are as follows (T is treatment, C is C. sativa and P is P. menziesii):
Fig. 1 - Framework of the different planting designs of C. sativa (C) and P. menziesii (P) plots at the study site. The grey line separates the plots destroyed by a wildfire in 2003 (upper part) from those survived to fire.

(1) Mixtures
- Row mixtures with species changing in the planting line:
- T1: 25% C. sativa 75% P. menziesii - 1 C and 3 P in the line (25C:75P)
- T2: 25% C. sativa 75% P. menziesii - 2 P and 1 C in the line (25C:75P)
- T3: 50% C. sativa 50% P. menziesii - 1 C and 1 P in the line (50C:50P)
- Lines mixtures with species changing between planting line:
- T4: 50% C. sativa 50% P. menziesii - 1 line of C and 1 line of P (50C:50P)
- T5: 25% C. sativa 75% P. menziesii - 2 lines of P and 1 line of C (25C:75P)
(2) Pure treatments
- T6: 100% C. sativa (100C:0P)
- T7: 100% P. menziesii (0C:100P)
Details about the plantation establishment are documented in Maia ([39]) and Luís & Monteiro ([37]).
Field work and laboratory methods
After the establishment of the research site several basic biometric measurements were made to analyze tree growth in the different treatments ([37]). All trees were numbered in each block and the initial height and diameter at breast height (DBH - 1.30 m above the ground) of each tree were measured in 1988. The height and DBH were measured again in 1992, 1996, 1998, 2000 and 2008. The final assessment was made in 2009.
DBH was measured with a steel diameter tape and two measurements were made in each tree to avoid errors. These data were used to calculate the basal area and density. Total height, defined as the maximum vertical distance from the ground level to the tree top, was measured for all the trees using a height pole. Averages of these variables were calculated for all treatments.
Soil depth was measured at four sampling locations in each plot, and the average was calculated to represent the soil depth at the plot. Moreover, soil samples in two horizon layers (horizon A: 0-20 cm; horizon B: 21-60 cm depths) were collected to analyze soil properties ([46]) and to test if species composition had effect on soil nutrient content.
Data are available for all plots until the year 2000. In 2003 a wildfire destroyed all the seven treatments in block I and two plots of mixtures in block II. Since then, measurements were made only on plots saved from the fire.
Aboveground biomass and net primary production
Tab. 1 - Allometric coefficients for the species studied using the model reported in eqn. 1. (B): biomass component (dry biomass - Kg); (a) and (b): model parameters. R2 was calculated using the transformation: ln(B) = ln(a) b ln(DBH). Sources: Ruiz-Peinado et al. ([59]) for C. sativa; Gower et al. ([24]) for P. menziesii.
| Species | Compartment | a | b | R2 |
|---|---|---|---|---|
| Castanea sativa | Stem (wood bark) | 0.0878 | 2.1474 | 0.94 |
| Branch > 7cm | 0.0004 | 3.491 | 0.902 | |
| Branch > 2 < 7cm | 0.1008 | 1.9089 | 0.83 | |
| Branch < 2cm | 0.2118 | 1.6938 | 0.858 | |
| Foliage | 0.15 (BStem + BBranches) |
|||
| Pseudotsuga menziesii | Stem wood | 0.0294 | 2.798 | 0.986 |
| Stem bark | 0.017 | 2.414 | 0.968 | |
| Live branch | 0.0184 | 2.033 | 0.895 | |
| Dead branch | 0.0184 | 1.642 | 0.786 | |
| New twig | 0.0003 | 2.166 | 0.924 | |
| New foliage | 0.0021 | 1.966 | 0.886 | |
| Total foliage | 0.0419 | 1.754 | 0.922 | |
Total aboveground biomass was calculated for each tree component (stem, branch and foliage) using allometric equations in all the pure and mixed treatments (Tab. 1). Biomass components of both species were estimated using the following model (eqn. 1):
where B is the biomass component (oven-dried weight - Kg), DBH is the diameter at breast height (cm) and a and b are the model parameters. Total biomass was calculated as the sum of the dry biomass of all components: tree leaves/needles, branches and stem. The allometric equations used to estimate biomass for C. sativa were taken from Ruiz-Peinado et al. ([59]), wherein tree data were collected in the Central Mountain Range and Sierra de Ronda, Spain. Patrício ([49]) developed specific equations to predict tree biomass per plant component of C. sativa high forest stands located in the northwest of Portugal. However, these equations were not considered in this investigation because of relevant differences between the two datasets as for the range of tree size and age. For P. menziesii biomass equations were taken from a work by Gower et al. ([24]) carried out in the Cascade Range of central Washington State (USA), given that no specific allometric studies are available for this species in Portugal. For both species, the range of tree structural characteristics corresponds to that observed in our study area.
Aboveground net primary production (ANPP) was calculated for all trees on each of the 512 m2 plots as the sum of average woody biomass increment (ΔB), of stem and branch, and the annual foliage biomass production. Herbivory was not estimated because it is very difficult to be assessed. However, previous studies suggest it to be less than 10-15% of the net primary production for forests ([25]). Biomass increments were calculated for each tree component for 2008 and 2009, based on annual diameter increment data and using allometric equations for each species.
Aboveground litterfall was estimated using 40 x 60 cm litter screens randomly placed inside each plot. Litter screens were deployed in July 2008 and litter was collected every 3 months for one year. Litter samples were dried at 70 °C to a constant mass and weighed (± 0.01 g) to determine the dry biomass.
The productivity results in pure and mixed treatments were then compared to provide information on which type of treatment had higher productivity.
Leaf area index
Leaf area index is defined as the projected area of tree foliage in relation to the total stand area. Leaf area index was calculated as the product of leaf biomass times the specific leaf area. Specific leaf area (fresh area to dry mass, cm2 g-1) per species was calculated from 125 leaves/needles randomly chosen from all canopy positions in 5 trees of each species in each treatment. Samples were collected from both species in the mixtures. All leaves were collected in the summer of 2011 after full leaf expansion and before the fall of the deciduous leaves. The projected leaf area of fresh leaf samples was measured in laboratory using a digital image analyzer for each scanned leaf. Reference objects of known area were used for calibration purposes. All samples were oven-dried at 70 °C to a constant dry mass. Specific leaf area was calculated as the sum of the leaf area divided by the sum of their dry mass.
Data analysis
Analysis of variance of a randomized complete block design were performed using the twenty-one plots organized in three blocks with four proportions according to the species composition: pure C. sativa, pure P. menziesii, mixtures with 1:1 and 1:3 proportions. The equality of means in DBH, height, the height to diameter ratio and aboveground biomass and ANPP among the four proportions were tested. Specific leaf area and leaf area index were also analyzed. Normality and variance homogeneity of residuals were verified. Student-Newman-Keuls’s (S-N-K) test was used to determine significant differences within species proportions in balanced and unbalanced design. All statistical analyses were performed using the package IBM SPSS Statistics® (version 21).
The yield of pure and mixed stands is usually compared on a relative basis; thus, the effects of combining the two species were evaluated by comparing the yield of each species in the mixture with its yield in monoculture as per Harper ([27]). To analyze the growth outcome of the mixed-species stands in this replacement series, a relative yield (RY) was calculated. The yield variable for this analysis was aboveground biomass per hectare, an adequate way to evaluate the productivity whenever the species have different basic densities ([44]). The RY of each species and the relative yield total (RYT) were calculated as follows (eqn. 2, eqn. 3, eqn. 4):
This index indicates the outcome of all interactions occurring in the stand, including both species interactions and the yield-density functions of either species.
If both species use resources in identical ways, i.e., compete for these resources, RY of each species will be equivalent to its proportional contribution in the mixture, with an expected RYT = 1. RY equal to the proportion of the given species in mixture indicates that, on average, trees were the same size in mixture or monoculture ([18]). An RYT > 1 indicates either niche separation or the existence of some beneficial relationship between species, leading to a potential productivity gain for the mixture. Contrastingly, RYT < 1 indicate an antagonistic or competitive relationship between the species in the mixture. In this study the assumption of the 1:1 mixture proportion grown independently would result in each species having an expected RY = 0.5 and RYT = 1.0.
Results
Patterns of growth
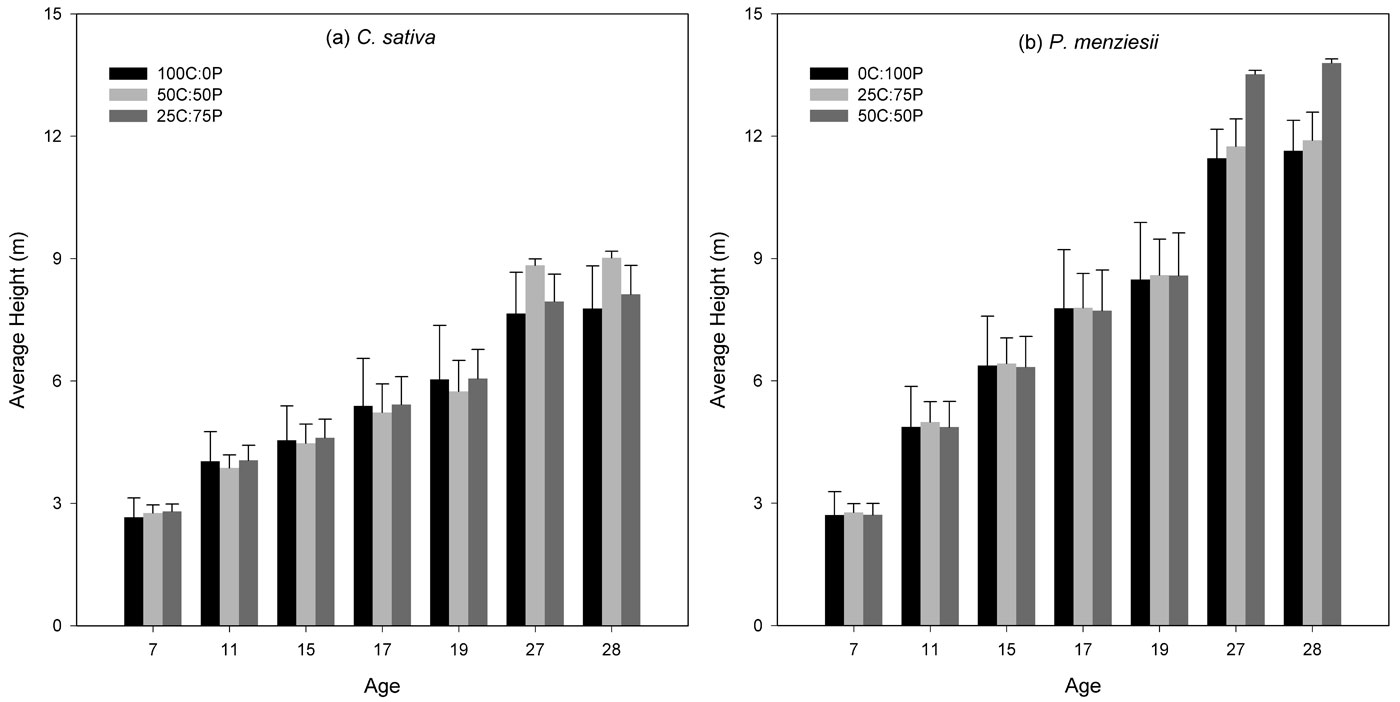
Tree height for C. sativa and for P. menziesii was not significantly different among the treatments for either species (P > 0.05) for all the studied years (Fig. 2). For C. sativa, tree height in different treatments was very similar early in the study, but as the study continued C. sativa in pure and 25C:75P treatments become taller until the age 19, when mixtures reached the higher average height (Fig. 2a). C. sativa had a smaller growth in pure treatments than in mixtures, on average 0.15 and 0.26 m less in the 1:3 and 1:1 proportions, respectively. Tree height for P. menziesii was also very similar among treatments early in the study, but at age 27 and 28, an increase in height growth in mixtures was observed; the 25C:75P and 50C:50P treatments have an average height with 11.9 ± 0.4 and 13.8 ± 0.1 m, respectively (Fig. 2b). Comparing the two species revealed that P. menziesii was taller than C. sativa (by more than 2 m at age 19 and 4 m at age 28) in all treatments.
Fig. 2 - Mean total height over time by species for the studied years. Height values were not significantly different (P > 0.05) among treatments for both C. sativa and P. menziesii.
Average DBH for all the treatments and species is displayed in Fig. 3. Since the start of the study (1988) DBH was not significantly different among treatments for both species (P > 0.05), although significant differences were observed later in the study for P. menziesii. C. sativa presented the higher DBH growth in mixtures compared with the pure treatment. The treatment 25C:75P had the higher DBH in the first 19 years, with 9.5 ± 2.3 cm in 2000 (Fig. 3a), and at age 28 this tendency was maintained, with an average DBH of 13.2 ± 1.4 cm in the treatment 25C:75P. P. menziesii 50C:50P treatments had higher DBH values compared with the pure and the 25C:75P treatments since age 11 with no significant differences (P > 0.05), although at age 27 and 28 the DBH in 50C:50P treatment was significantly different (P < 0.05) from the pure treatment and 1:3 proportions. These results may indicate that radial growth of P. menziesii is higher when mixed with C. sativa.
Fig. 3 - Mean diameter at breast height (DBH) over time by species for the studied years. DBH values were not significantly different among treatments for C. sativa (P > 0.05), but were significantly different for P. menziesii. Differences in letters above the bars indicate significant differences in DBH between treatments (P < 0.05).
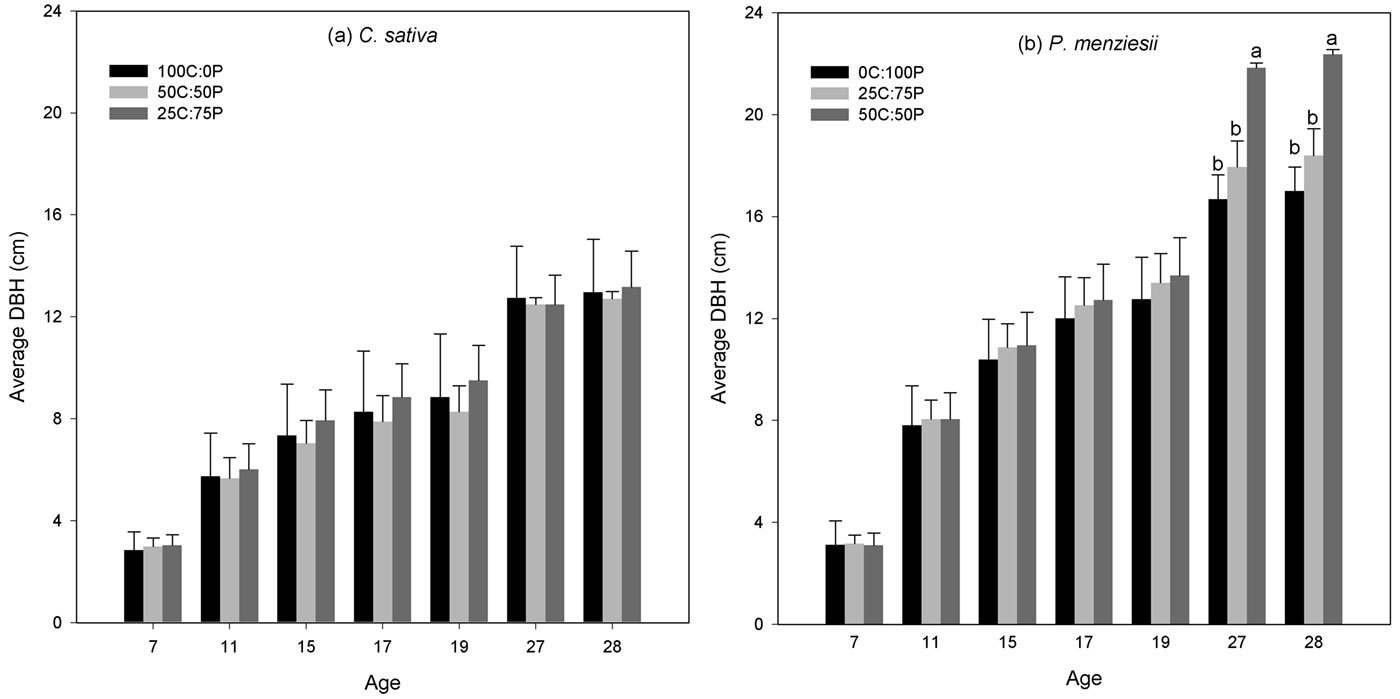
Results show that P. menziesii was thicker than C. sativa (by more than 4 cm at age 19) in all treatments. Furthermore, these differences appeared early in stand development and were already in the order of 2 cm at age 11.
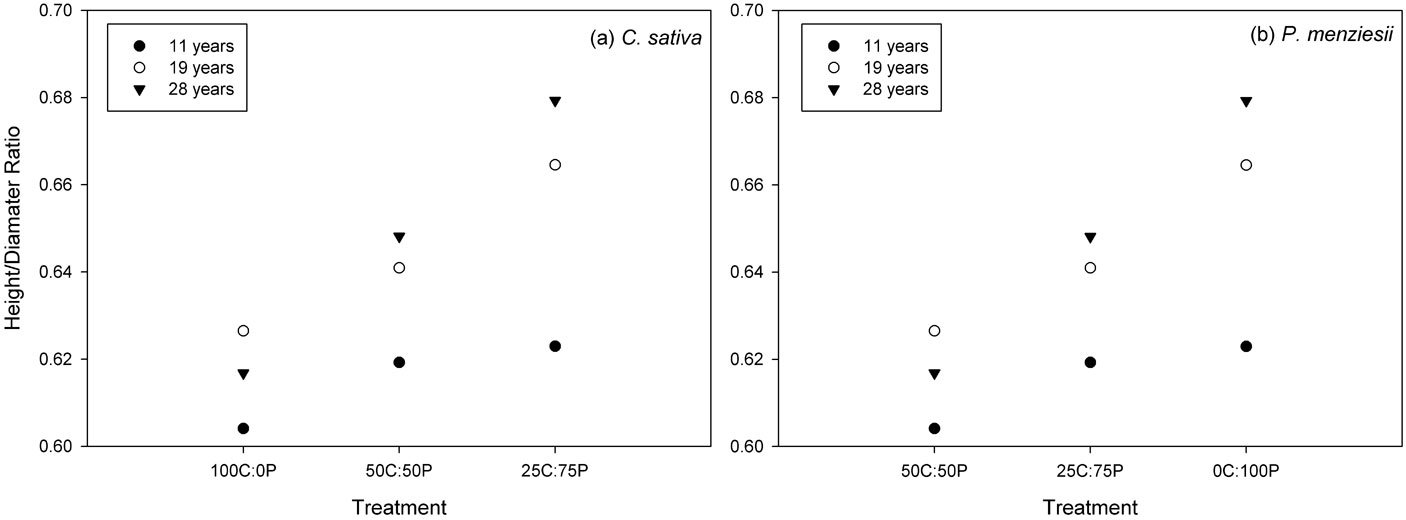
The height to diameter at breast height ratio (h/d) was compared among species growing alone and together at age 11, 19 and 28 (Fig. 4). Changes in the h/d patterns may reflect competition between species. Even though some differences in diameter and height growth were evident over the period considered, the h/d ratio was very low. C. sativa growing in mixtures had a slightly greater h/d ratio than in pure treatments, but no significant differences were observed among the treatments (P > 0.05). P. menziesii instead had slightly greater h/d ratio when growing in pure treatments. Differences among treatments were statistically significant (P < 0.05) only at age 28.
Fig. 4 - Height/diameter ratio (h/d) by treatment at age 11, 19 and 28. Significant differences (P < 0.05) in h/d ratio among treatments are indicated by different letters.
The effects of combining the two species considered in a mixture were analyzed by comparing the yield of each species in mixture with its yield in a pure stand ([27]). Relative yield (RY) and relative yield total (RYT) were calculated based on the aboveground biomass per hectare for both proportions of mixtures at all the studied ages (Fig. 5).
Fig. 5 - Relative yield (a) and relative yield total (b) of aboveground biomass per hectare between age 7 and 28 for C. sativa and P. menziesii in 1:1 and 1:3 mixtures.
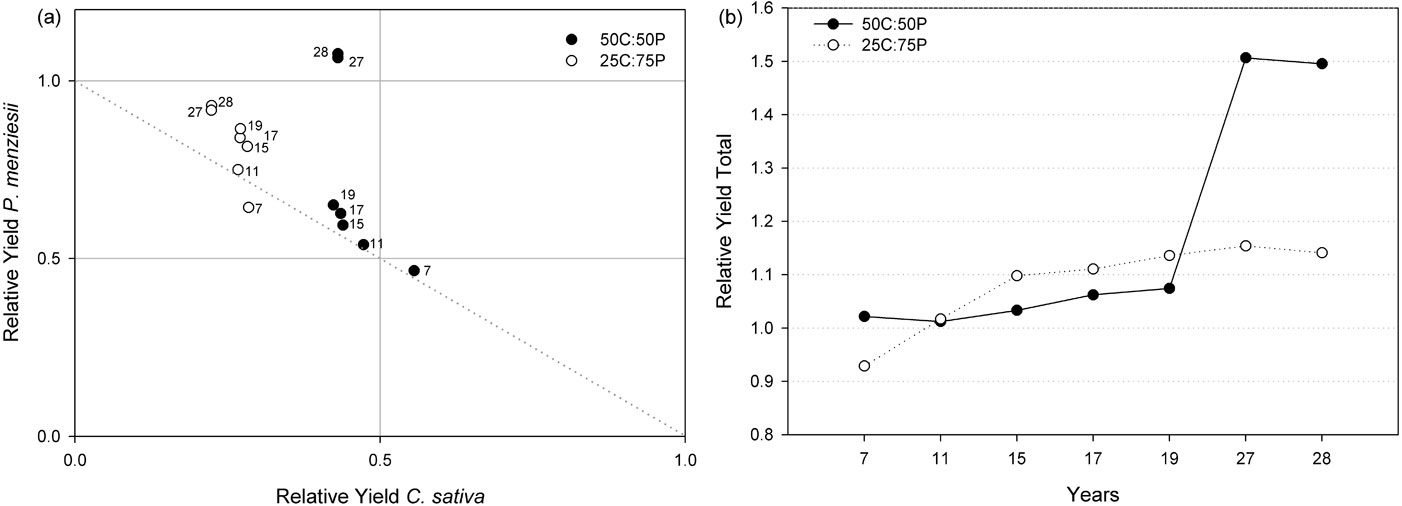
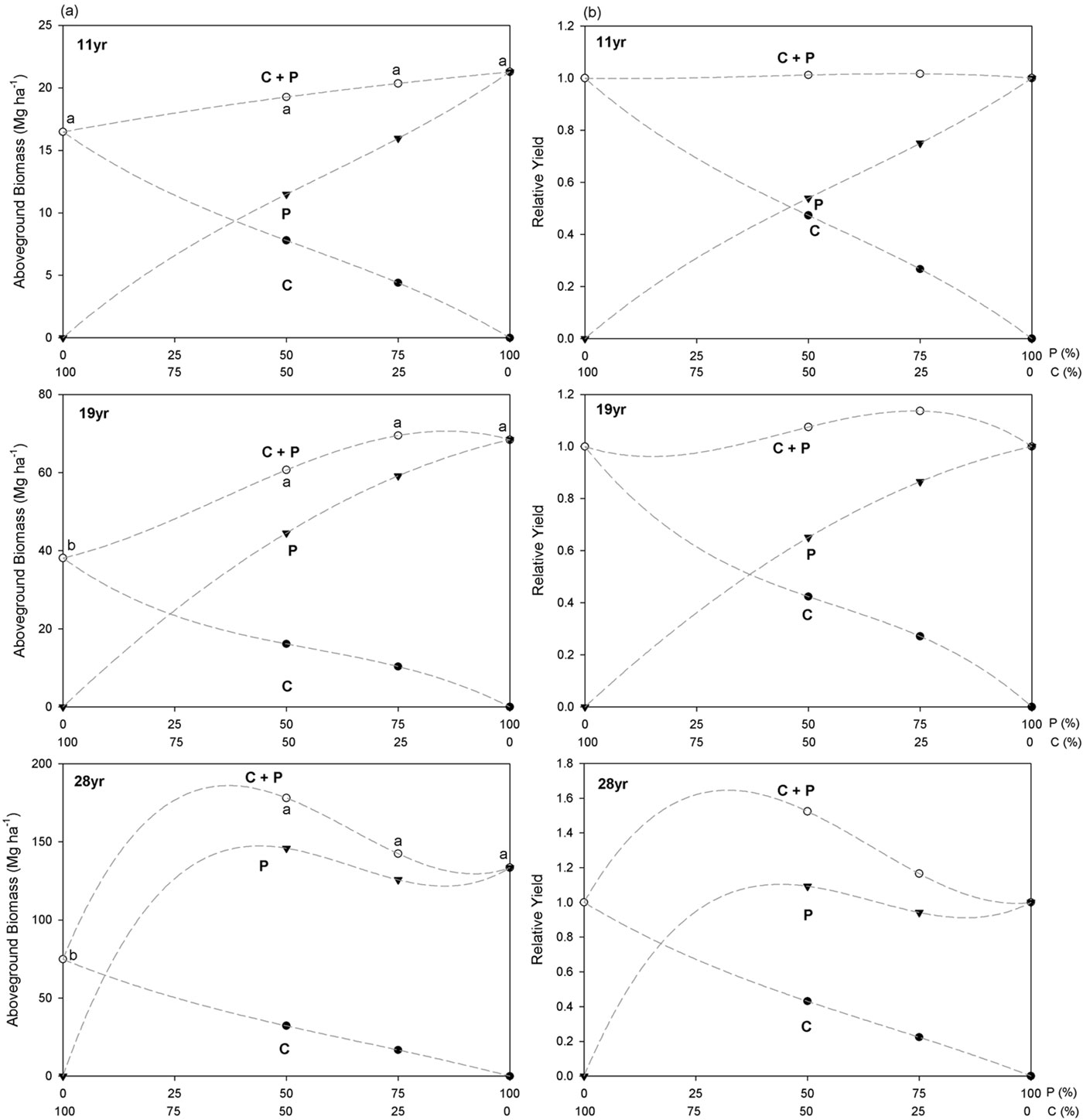
For the 1:1 mixture (50C:50P), the RY index for C. sativa was less than 0.5 since age 11 (except for age 7). P. menziesii had RY values substantially greater than 0.5 since age 11, and at age 27 and 28 the RY was greater than 1.0. It is clear that the mixture of the two species benefited the P. menziesii since age 11, and this species had the competitive advantage over C. sativa on this proportion. Plotting together the RY index obtained for both species, all the values were above 0.5 (above the dashed line reported in Fig. 5a), indicating a higher productivity in mixture. Combined RYT was higher than 1.0 for all the studied years (Fig. 5b). Thus, in this proportion, it appears that significant niche separation between these species may exist.
For the 1:3 mixture (25C:75P), the RY of C. sativa was greater than 0.25 until age 20, and slightly lower afterward. P. menziesii instead, had RY values smaller than 0.75 until age 11 and greater since age 15. RYT for this proportion was higher than 1.0 only after 11 years since planting (Fig. 5b).
Fig. 6b combines the results obtained for absolute and relative yield at 3 ages (11, 19 and 28 since planting) for the two proportions of mixtures considered. At the age 11 both the relative and absolute yield (aboveground biomass per hectare) for the mixtures were indistinguishable from those of monocultures. Contrastingly, differences between mixtures and monocoltures in both relative and absolute yield are fairly evident at age 19 and 28.
Fig. 6 - Aboveground biomass (a) and absolute and relative yield of aboveground biomass per hectare (b) for the C. sativa (C) and P. menziesii (P) species at age 11, 19 and 28, with the total number of individuals constant. The proportion of each species in mixture, per treatment, changes from no individuals (0%) up to the total number of individuals (100%). Significant differences (P < 0.05) for aboveground biomass among treatments are indicated by different letters.
Plot characteristics
Specific leaf area was not significantly different between treatments in P. menziesii, but it was significantly different (P < 0.05) in the C. sativa 50C:50P treatment when compared with the pure treatments (Tab. 2). Pure treatments of C. sativa showed average specific leaf area values nearly doubled compared to pure P. menziesii, with values of 122.0 ± 5.7 cm2 g-1 and 63.5 ± 2.3 cm2 g-1, respectively. Average values of mixtures reached higher values compared to the pure treatments. The 50C:50P treatments had the higher values in both species; the highest value was for C. sativa.
Tab. 2 - Specific leaf area (fresh area: dry mass) by species. Different letters indicates significant differences among treatment means (P < 0.05).
| Species | Plot | Specific leaf area (cm2 g-1) | ||
|---|---|---|---|---|
| Mean | Range | Standard error |
||
| C. sativa | 100C:0P | 122 b | 95.2-144.9 | 5.7 |
| 50C:50P | 172.8 a | 138.3-189.9 | 6.4 | |
| 25C:75P | 143.1 ab | 112.2-204.5 | 9.6 | |
| P. menziesii | 0C:100P | 63.5 a | 53.8-75.6 | 2.3 |
| 25C:75P | 57.2 a | 44.9-78.1 | 3.2 | |
| 50C:50P | 67.2 a | 55.2-89.6 | 4.3 | |
Mean stand characteristics at age 28 for the two pure treatments and mixtures are summarized in Tab. 3. At this stage, the C. sativa pure treatment presented a DBH average of 13.0 ± 2.5 cm (range: 4.5-22.4 cm) and a basal area of 14.7 ± 5.9 m2 ha-1. The average height ranged from 3.3 to 10.8 m. The P. menziesii pure treatment presented higher values of DBH, basal area and height compared to pure C. sativa. As for density, the pure treatments had similar values, 1064 trees ha-1 for C. sativa and 1104 trees ha-1 for P. menziesii, which represented 85% and 88% of the total trees planted, respectively. Mean DBH, height and basal area for P. menziesii was 17.0 ± 1.0 m (range: 4.2-26.3 m), 11.6 ± 0.9 m (range: 4.1-16.1 m) and 25.4 ± 5.4 m2 ha-1, respectively. The mixtures presented some heterogeneity. P. menziesii DBH and height were higher in the 50C:50P treatment, but C. sativa higher values of DBH were observed in the 25C:75P treatment and higher in height in the 50C:50P treatment. Basal area of C. sativa in 50C:50P was less than half of the pure C. sativa treatment, and less than expected in the 25C:75P, indicating a negative effect on C. sativa growing in mixtures. Total LAI was not significantly different across treatments (P > 0.05), ranging from 2.3 m2 m-2 for 0C:100P to 5.9 m2 m-2 for 100C:0P. While the contribution of P. menziesii to the total value of LAI in each plot decreased by 0.5 m2 m-2 from the pure to the mixed treatments, the contribution of C. sativa LAI decreased by 4.0 m2 m-2.
Tab. 3 - Structural characteristics at age 28 by species composition. Mean values are reported (standard error in parentheses). (DBH): average diameter at breast height; (H): average tree height; (N): number of trees per hectare; (BA): average basal area; (SD): average soil depth; (LAI): average leaf area index; (*): Mean values with different letter within LAI differ significantly among the treatments (P < 0.05).
| Treatment | Species | DBH (cm) |
Range DBH (cm) |
H (m) |
Range H (m) |
N | BA (m2 ha-1) |
SD (cm) |
LAI (m2 m-2) |
|---|---|---|---|---|---|---|---|---|---|
| 100C:0P | C. sativa | 13.0 (2.5) | 4.5 (2.1) - 22.4 (3.4) | 7.8 (1.3) | 3.3 (0.9) - 10.8 (1.3) | 1064 (29) | 14.7 (5.9) | 77 (3.9) | 5.9 (2.4) a |
| 50C:50P | C. sativa | 12.7 (0.5) | 5.6 (0.2) - 19.2 (0.2) | 9.0 (0.3) | 4.7 (1.3) - 12.4 (0.3) | 508 (39) | 6.4 (0.0) | 74 (2.2) | 5.7 (0.2) a |
| P. menziesii | 22.4 (0.3) | 6.4 (1.8) - 34.4 (1.5) | 13.8 (0.2) | 4.7 (1.4) - 18.6 (0.1) | 596 (29) | 23.4 (1.9) | |||
| 25C:75P | C. sativa | 13.2 (1.7) | 6.0 (0.9) - 20.3 (1.6) | 8.1 (0.9) | 4.5 (0.7) - 11.1 (0.9) | 244 (19) | 3.3 (0.6) | 70 (2.9) | 3.3 (0.4) a |
| P. menziesii | 18.4 (1.3) | 6.8 (1.3) - 28.9 (2.0) | 11.9 (0.9) | 6.0 (0.7) - 16.5 (0.6) | 827 (27) | 22.3 (2.9) | |||
| 0C:100P | P. menziesii | 17.0 (1.0) | 4.2 (0.1) - 26.3 (0.9) | 11.6 (0.9) | 4.1 (0.5) - 16.1 (0.6) | 1104 (88) | 25.4 (5.4) | 75 (2.4) | 2.3 (0.5) a |
Total biomass and aboveground net primary production
In order to analyze the growth dynamics of the species, the aboveground biomass was estimated at the age 11, 19 and 28, i.e., in the beginning, the middle and the last year of available field data. P. menziesii pure treatments had higher aboveground biomass compared to C. sativa pure treatments at the 3 ages analyzed, with 133.4 ± 30.8 Mg ha-1 for P. menziesii and 74.8 ± 30.6 Mg ha-1 for C. sativa at age 28. At age 11 (Fig. 6a, 11yr) results reveal a low interaction between species (intra- and interspecific competition are the same), since the aboveground biomass in mixtures is similar to pure treatments with no significant differences (P > 0.05) between treatments. The growth of the two species in mixture is the result of the contribution of each species to the total aboveground biomass in direct ratio to its original proportion. However, after only eight years, biomass dynamics were greatly influenced by the planting design for the two species, and P. menziesii contributed positively to the overall positive mixing effect in both 50C:50P and 25C:75P treatments. The interspecific competition was higher than the intraspecific competition. P. menziesii had a greater share of resources in mixture than in pure treatments. Aboveground biomass of C. sativa in 50C:50P was less than half of the pure C. sativa treatment, and a similar pattern was found for the 25C:75P treatment. The C. sativa pure treatment had lower biomass with significant differences (P < 0.05) from the other treatments (Fig. 6a, 19yr). The differences in aboveground biomass continued to increase until 2009, at age 28. After this age, the experimental design was unbalanced because of the wildfire occurred in 2003. Nonetheless, the post-hoc tests carried out on data collected in 2009 (and 2008) confirmed the results obtained for previous years (1998 and 2000), when the design was complete and fully balanced. The aboveground biomass in mixtures was significantly different from pure C. sativa treatments (P < 0.05), but no significant differences from pure P. menziesii treatments were found. The 100C:0P treatment still had the lowest aboveground biomass. These results indicated greater aboveground biomass in pure P. menziesii and in mixtures than in pure C. sativa treatments. Looking only at mixed treatments, the higher value of aboveground biomass was observed in treatments with a higher proportion of deciduous species (Fig. 6a, 28yr).
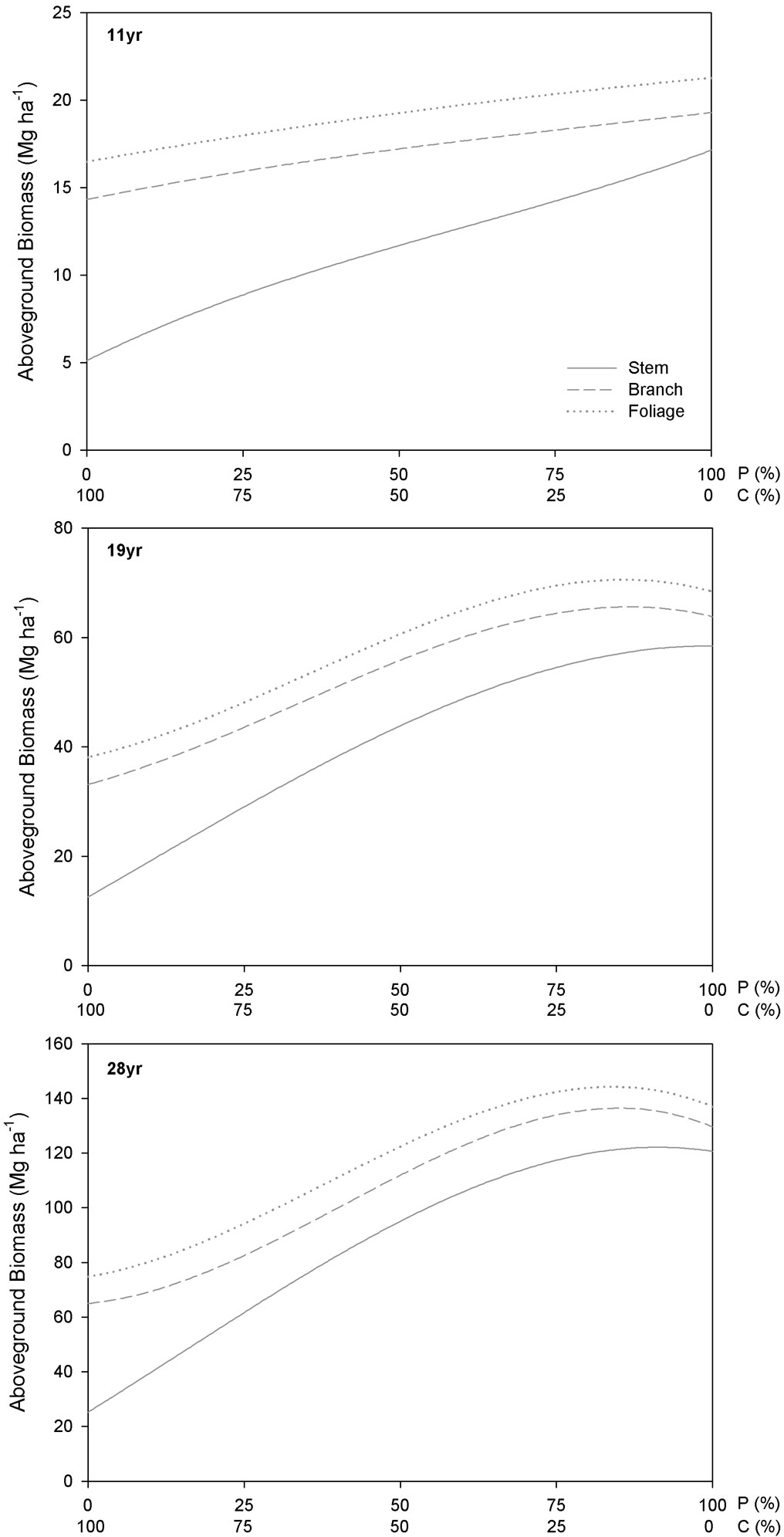
Analysis of the aboveground biomass per tree component among treatments at age 11 indicated larger stem biomass for P. menziesii (17.1 ± 9.4 Mg ha-1) and greater branch and foliage biomass for C. sativa pure treatments (9.2 ± 4.6 and 2.2 ± 1.1 Mg ha-1, respectively - Fig. 7, 11yr). Similar results were found for biomass components at age 19 (Fig. 7, 19yr). However at age 28, stem biomass was higher in the 50C:50P treatment (142.3 ± 11.3 Mg ha-1, 80% of the total biomass) in respect to other treatments, and the foliage biomass with 10.5 ± 0.5 Mg ha-1 was also higher (Fig. 7, 28yr). Branch biomass was greater in the 100C:0P treatment (39.8 ± 15.9 Mg ha-1, 53% of the total biomass; stem biomass comprised 32% of the total biomass).
Fig. 7 - Aboveground biomass per hectare and tree component (values of branches and foliage are cumulated to the stem values) for the C. sativa (C) and P. menziesii (P) species at age 11, 19 and 28, with the total number of individuals constant. The proportion of each species in mixture, per treatment, changes from no individuals (0%) up to the total number of individuals (100%).
ANPP (total and by tree component) was calculated for each treatment for 2008 and 2009 (Tab. 4). Results from the total ANPP analysis indicated higher productivity in mixtures compared with pure treatments, although no significant differences (P > 0.05) were found. In respect to pure treatments, C. sativa had a higher productivity (7.9 ± 0.0 Mg ha-1 yr-1, 18% and 70% of the total ANPP due to the branch and foliage components, respectively) than P. menziesii, the latter being the treatment with the lowest ANPP value (7.0 ± 0.8 Mg ha-1 yr-1, with 84% of the total ANPP due to the stem component). Finally, significant differences among treatments (P < 0.05) were found in branch, stem and foliage components.
Tab. 4 - Aboveground net primary production (Mg ha-1 yr-1), total and per tree component at age 28 (percentage of total ANPP in each component in parentheses). Values ± standard error in the same column followed by the same letter are not significantly different (P > 0.05).
| Treatment | Stem | Branch | Foliage | Total |
|---|---|---|---|---|
| 100C:0P | 0.93 ± 0.5 b (12%) |
1.39 ± 0.7 a (18%) |
5.55 ± 1.2 a (70%) |
7.86 ± 0.0 a |
| 50C:50P | 8.93 ± 0.7 a (61%) |
0.95 ± 0.1 ab (6%) |
4.87 ± 0.7 ab (33%) |
14.76 ± 1.6 a |
| 25C:75P | 7.14 ± 1.5 a (66%) |
0.63 ± 0.1 ab (6%) |
3.02 ± 0.4 ab (28%) |
10.78 ± 1.9 a |
| 0C:100P | 5.83 ± 0.7 ab (84%) |
0.31 ± 0.0 b (4%) |
0.84 ± 0.2 b (12%) |
6.98 ± 0.8 a |
Discussion
Growth patterns
Stratified canopies in mixed stands tend to develop naturally because of differences in the auto-ecology of the species involved. Shade-intolerant species generally have higher rates of juvenile height growth than shade-tolerant species, while shade-tolerant species are able to survive in reduced light environments ([30], [47], [60]). Kelty ([31]) also stated that the overall growth rate of mixed-species plantations with stratified canopies would be affected by their shade tolerance as well as the density of the upper canopy.
In this study the influence of mixtures on the height of both species was evident all along the analyzed timespan. P. menziesii and C. sativa differ in their tolerance to shade, and different height growth patterns (along with stratification) were observed for the two species after 11 years of planting. Interspecific competition on mixtures also leads to stratified canopies ([34]). In this study, C. sativa formed the lower canopy stratum in the mixture as a consequence of its lowered height growth. Amoroso & Turnblom ([3]) found greater height, diameter and individual tree volume for P. menziesii than for Tsuga heterophylla (Rag.) Sarg. in 50/50 mixture plantations. Analogously, P. menziesii showed a higher total height in this study, making up the upper part of the canopy in the mixtures. Other studies also found greater yield in mixed than in pure stands for most tree components ([29], [10], [13], [32], [41], [4], [21]). This study shows that by age 19 P. menziesii has outgrown C. sativa by 4 m (on average). Differences in growth between the two species had been increasing over the period of time measured, and are expected to continue in the near future.
The observed difference between C. sativa DBH growing in pure and 50C:50P treatments was at most 0.7 cm. Such small difference indicates that C. sativa growth was not significantly influenced by the presence of P. menziesii. Nevertheless, a higher mean value was observed in the 25C:75P treatment compared with the pure plots, though no significant differences among treatments were observed. Mean DBH of P. menziesii amounted to 22.4 cm in the 1:1 proportion, compared to 17.0 cm in the pure treatments (+31%, P < 0.05). In contrast, C. sativa achieved an average DBH of 12.7 cm in the 1:1 proportion and 13.0 cm in the pure treatments. Pretzsch et al. ([51]) observed similar behavior in pure and mixed stands of Norway spruce (Picea abies (L.) H. Karst.) and European beech (Fagus sylvatica L.) throughout central Europe.
Dominant trees within a stand tend to capture more light and to use it more efficiently for their growth. According to Binkley et al. ([9]) faster growth of large trees typically results from a combination of increased light absorption (accounting for about three-fourths of the effect) and increased efficiency of light use (about one-fourth of the effect). Light use and light use efficiency were not analyzed in the present study, although results from Campoe et al. ([12]), Gspaltl et al. ([26]) and Le Maire et al. ([35]) provide new information and understanding about growth rates. Campoe et al. ([12]) found in a study with Eucalyptus grandis that the faster growth of dominant trees was driven exclusively by a more efficient light use. Contrastingly, Gspaltl et al. ([26]) observed that larger trees used more light and use the intercepted light more efficiently for growth in a study on Picea abies, while Le Maire et al. ([35]) observed a more efficient light use of taller trees in Acacia mangium and Eucalyptus grandis. Hints of a higher light use efficiency for dominant trees are also strong in these studies. Future works on how light use efficiency accounts for differences in growth among C. sativa and P. menziesii will contribute for a better understanding of the mechanisms involved in overyielding in mixtures.
Interspecific and intraspecific competition
The more efficient utilization of site resources by different species in mixed stands may result in a greater total productivity in comparison to pure stands, as a consequence of the less intense interspecific than intraspecific competition in mixed stands ([30]).
In this study, interspecific and intraspecific competition were assessed by comparing tree growth in mixture and in pure treatments for each species ([42]). After 20 years of planting, P. menziesii experienced a substantial increase in DBH, height, and aboveground biomass in mixtures compared to their growth in pure treatments. The analysis of height/diameter ratio (h/d) can give an insight on interspecific and intraspecific competition ([33]). Trees allocate more carbon to height than to diameter growth to participate in the canopy ([4]). The slightly lower h/d that P. menziesii trees exhibited in mixtures compared to monoculture is consistent with the results by Wang et al. ([62]), who reported that shade-intolerant species growing in mixed stands allocated more carbon to stem wood. C. sativa, instead, presented higher (though not significant) h/d ratios in mixtures compared to the pure treatments. This result suggests that interspecific competition is greater than intraspecific competition in the mixed stands studied, in that C. sativa needed to allocate relatively more resources to height growth to participate in the canopy, while P. menziesii reach the canopy earlier and could therefore allocate more resources to its radial growth. However, our results confirmed that the development of mixed-species plantations is highly influenced by the relative growth rate of each species ([16]). When P. menziesii was surrounded by a larger proportion of C. sativa, P. menziesii height and stem diameter was increased. Furthermore, interspecific competition between species influenced mainly the height and less the DBH in C. sativa, while for P. menziesii both height and DBH growth were affected.
The high productive potential of P. menziesii in the mountains areas of northern Portugal has been recognized ([36], [38]). Indeed, the good performances of this species in the study area may be explained by several environmental factors of the study site ([14]). However, Maia et al. ([40]) demonstrated that P. menziesii in Portugal performed better in terms of height growth than several other species, like Pinus pinaster Aiton, C. sativa, P. nigra var. maritima (Ait.) Melv., Cedrus atlantica (Endl.) Carr. and P. sylvestris L.
The small number of mixed-species plantations worldwide make difficult to identify the general patterns for tree interspecific interactions ([57], [18]), and to predict their effect all along the stand development ([15]). Nonetheless, evaluation of interaction and competition between neighboring trees in this study is still being done.
Overall productivity: pure vs. mixed stands
The productivity between pure and mixed treatments was compared both on a relative (relative yield, sensu [27]) and absolute basis (aboveground biomass per ha). In mixture 1:1, the RYT was higher than 1.0 for all the studied years. Since age 11, the relative yield for P. menziesii was as expected (0.5) or higher than expected; instead, it was lower than expectations for C. sativa. In the mixture 1:3, a combined RYT value higher than 1.0 was also found, except at age 7. This could imply a nursing effect by the species in mixtures. Bauhus et al. ([4]) observed similar RY in a mixed plantation.
Aboveground biomass per ha had similar yield in pure and mixed treatments in the first years after plantation. As the years passed, an increase in aboveground biomass was observed in mixtures (in comparison to pure plots) as a result of the enhanced biomass of P. menziesii, probably due to an increase in light interception ([3]). In contrast, the yield of C. sativa was lower compared with the results in pure treatments. This may indicate a positive interaction and a complementary benefit between C. sativa and P. menziesii when growing together ([15]). According to Kelty ([31]), the competition intensity depends on the ability of mixtures to better exploit site resources for biomass production compared to pure treatments. Canopy stratification of both species ([15]) can also partially explain these results, as well as a better use of the site resources ([3]). The multilayer structure with light gaps enables subdominant and understory C. sativa trees to survive in the plantation. Amoroso & Turnblom ([3]) observed similar results with an increase in productivity in mixtures with high densities. Species interactions involve a complex balance of competition and facilitation ([30], [11], [51]). Competition and facilitation are frequently coupled, making them difficult to distinguish experimentally ([11]).
Higher productivity in mixtures indicates a facilitation of P. menziesii by C. sativa, since the productivity of the former in mixtures exceeds the productivity in pure treatments. ANPP variations in pure C. sativa treatments where consistent with the results from Zianis & Mencuccini ([63]), although the stem wood production value was lower than the one observed by the above authors. Stem wood production depends not only on total net production but also on its allocation ([63]), and stem has a longer residence time of cumulative biomass than both branches and foliage. On average, in pure C. sativa treatment, 88% of ANPP was allocated to crown (branch and foliage), whereas in pure P. menziesii and mixed treatments the allocation was about 16% and up to 33%, respectively. Binkley et al. ([9]) reported that stands with low basal area typically have less stem wood than stands with moderate or high basal areas, largely owing to smaller canopies and lower absorption of light. In this study, low basal area was found for the C. sativa pure treatment, where the stem component represents only 12% of the ANPP. The mixtures had higher (though not significant) average productivity values than the pure treatments. Considering that P. menziesii is a fast growing species providing revenues in the short term, and C. sativa a more valuable species with slower growth and revenues in the long term, it will be important to monitor species growth and evaluate their productivity to the final rotation.
Conclusion
C. sativa and P. menziesii growth and productivity were assessed in mixtures and pure treatments within the same planting site, under shared soil and climatic conditions. Overall, mixtures reached higher yield than pure treatments. Moreover, positive interactions in mixtures have been found between C. sativa and P. menziesii in terms of aboveground productivity, leading to higher ANPP compared with pure treatments. Canopy stratification seems the major underlying mechanism (C. sativa in the lower and P. menziesii in the upper canopy stratum), with an increase in the light capture in mixtures. The results of this study improved our knowledge of the growth dynamics of these species in northern Portugal.
Acknowledgments
We are most grateful to Carlos Fernandes and Carlos Brito for their help with the field data collection, to Armindo Teixeira and Helena Ferreira for their help in the laboratory. This study was funded by FCT research in project PTDC/AGR-CF/68186/2006 and FCT Program SFRH/PROTEC/50127/2009.
References
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Escola Superior Agrária, Instituto Politécnico de Viseu (ISPV), Estrada de Nelas, Quinta da Alagoa, 3500-606 Viseu (Portugal)
CITAB, Universidade de Trás-os-Montes e Alto Douro (UTAD), Apartado 1013, 5000-911 Vila Real (Portugal)
CEABN, Instituto Superior de Agronomia, Tapada da Ajuda, 1349-017 Lisboa (Portugal)
Department of Forestry, 1630 Linden Drive, University of Wisconsin, Madison, 53706 Wisconsin (USA)
CIMO, Instituto Politécnico de Bragança (IPB), Escola Superior Agrária, Bragança (Portugal)
Corresponding author
Paper Info
Citation
Nunes L, Gower ST, Monteiro ML, Lopes D, Rego FC (2014). Growth dynamics and productivity of pure and mixed Castanea sativa Mill. and Pseudotsuga menziesii (Mirb.) Franco plantations in northern Portugal. iForest 7: 92-102. - doi: 10.3832/ifor1087-007
Academic Editor
Roberto Tognetti
Paper history
Received: Jul 26, 2013
Accepted: Sep 27, 2013
First online: Dec 18, 2013
Publication Date: Apr 02, 2014
Publication Time: 2.73 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2014
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 45047
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 37413
Abstract Page Views: 1836
PDF Downloads: 4271
Citation/Reference Downloads: 40
XML Downloads: 1487
Web Metrics
Days since publication: 3772
Overall contacts: 45047
Avg. contacts per week: 83.60
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Feb 2023)
Total number of cites (since 2014): 6
Average cites per year: 0.55
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Species interactions in pure and mixed-species stands of silver fir and European beech in Mediterranean mountains
vol. 14, pp. 1-11 (online: 02 January 2021)
Short Communications
Biomass production of young lodgepole pine (Pinus contorta var. latifolia) stands in Latvia
vol. 6, pp. 10-14 (online: 14 January 2013)
Research Articles
Influence of inorganic salts on biomass production, biochemical composition, and bioethanol production of Populus alba
vol. 13, pp. 566-574 (online: 07 December 2020)
Review Papers
Should the silviculture of Aleppo pine (Pinus halepensis Mill.) stands in northern Africa be oriented towards wood or seed and cone production? Diagnosis and current potentiality
vol. 12, pp. 297-305 (online: 27 May 2019)
Research Articles
Energy production of poplar clones and their energy use efficiency
vol. 7, pp. 150-155 (online: 23 January 2014)
Research Articles
Contrasted growth response of hybrid larch (Larix × marschlinsii), jack pine (Pinus banksiana) and white spruce (Picea glauca) to wood ash application in northwestern Quebec, Canada
vol. 14, pp. 155-165 (online: 06 April 2021)
Research Articles
Allometric equations to estimate above-ground biomass of small-diameter mixed tree species in secondary tropical forests
vol. 13, pp. 165-174 (online: 02 May 2020)
Research Articles
First vs. second rotation of a poplar short rotation coppice: leaf area development, light interception and radiation use efficiency
vol. 8, pp. 565-573 (online: 27 April 2015)
Research Articles
The potential of using xylarium wood samples for wood density calculations: a comparison of approaches for volume measurement
vol. 4, pp. 150-159 (online: 11 August 2011)
Research Articles
Variation of wood and bark density and production in coppiced Eucalyptus globulus trees in a second rotation
vol. 9, pp. 270-275 (online: 08 September 2015)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







