Field performance of poplar for bioenergy in southern Europe after two coppicing rotations: effects of clone and planting density
iForest - Biogeosciences and Forestry, Volume 5, Issue 5, Pages 224-229 (2012)
doi: https://doi.org/10.3832/ifor0628-005
Published: Oct 02, 2012 - Copyright © 2012 SISEF
Research Articles
Abstract
Short Rotation Forestry (SRF) plantations are estimated around 7 000 hectares in Italy, mostly established in northern regions. Recently thousands hectares of SRF were established in central and southern regions of Italy also (especially in Latium and Molise) due to the promising introduction of new clones resistant to summer water limiting and pest attacks. A poplar SRF plantation was established in central Italy in order to investigate the influence of site conditions and planting densities/spacings on the growth and yield production of three hybrid poplar clones (AF2, AF6 and Monviso). Cuttings of each clone were planted at two different densities/spacings and planting design as: (a) low density = 7 140 cuttings ha-1 in a single-row design (SR); (b) high density= 10 360 cuttings ha-1 in a twin-rows design (TR). Growth (both diameter and height) and yield were evaluated after biennial (2006-2007) and triennial (2008-2010) rotation. Relationships between fresh weight and diameter were determined to calculate the yield of each clone expressed in MgDM ha-1 years-1. At the end of the triennial rotation (2010) all poplar clones showed very high stump survival rates with higher values in SR (89.9%) compared to TR (85.6%). AF2 clone can be advisable thanks to the higher yield production than both AF6 and Monviso. Considering planting densities/spacings, SR seems to be more advisable than TR in order to obtain a good biomass production and a low planting costs. Considering the coppicing cycles, triennial rotation allows higher annual yield production (average value of 13.0 MgDM ha-1) than biennial rotation (average: 10.1 MgDM ha-1).
Keywords
Short Rotation Forestry, Hybrid Populus Clones, Coppicing Cycles, Yield, Growth, Optimal Density
Introduction
The international discussion on the climate change and the hypothesis that it is determined also by the human activities fosters the renewable energy uses. Renewable energy uses play a key role to honor the commitments that Italy has signed with the European Commission in order to achieve by 2020 for the whole Europe: (i) a reduction of 20% in energy consumption; (ii) a reduction of 20% in CO2 emission compared to 1990; (iii) more than 20% of the total energy demand will come from renewable energy. The Short Rotation Forestry (SRF) is one of the measures indicated by the article 3.4 (additional activities) of the Kyoto Protocol ([33]); they serve as feedstock for the pulp and paper industry but also can be established specifically to address the feedstock needs for energy or biofuel industry and to combat desertification processes. Among these, the woody biomass attracts great attention as a renewable energy and as alternative to fossil fuels (e.g., firewood, wood chips and pellets). The wood biomass production for energy has promising productive and environmental benefits, with a minor impact on food markets, and a significant potential to lower net CO2 emissions in the environment and improving the energy balance ([34], [17], [24]). SRF system is an intensive cultivation: the fast-growing hardwoods at high density are employed and the average period of rotation is less than 10 years ([30]). The Italian interest of SRF as a renewable energy began in the 1990s. Based on recent inventories ([6], [40]), the Italian surface of SRF was estimated around 7 000 hectares, mostly established in northern Italy, where the regional services of Lombardy, Veneto, Piedmont and Friuli released goods grants to establish and manage the SRF. Also European actions aimed to promote and extend the SRF since 2005 (Biomass Action Plan). Poplars (Populus spp.), especially modern hybrids, are well adapted to site conditions in Italy, and are the most common tree species used in SRF plantations ([26]) because they show great plasticity over a wide range of habitats. However, most regions in central Italy are affected by long summer dryness, resulting to be a suboptimal site for poplar cultivation, due to the very high water consumption of the woody crop in comparison with northern latitudes. For this reason some poplar clones were selected for growing in suboptimal sites of Italy.
Other species for SRF, such as willow (Salix spp.), black locust (Robinia pseudoacacia L.) and eucalyptus (Eucalyptus spp.) are still under research assessment and are not commercially available yet ([32]). This was because Robinia is strongly sensitive to water limiting; eucalypts are particularly sensitive to low winter temperatures; willows are also sensitive to water limitations. Indeed, they are currently used in others European country like Sweden ([23]), Hungary ([31]) and Portugal ([29]).
The SRF, owing to the high density and competition, grows with water and light limitations, associated with susceptibility to pest attacks ([15]). Hence, SRF plantations should be planted on “optimal sites”, with the support of “optimal cultural practices” to have higher productivity. Choosing the best planting density/spacing resulted to be a key-decision to ensure high production and high survival over time. This was because SRF plantations are characterized by continuous growth after their cutting, providing a living inventory of available biomass. Generally, harvesting of a SRF is carried out every two years (biennial rotation) over 5-6 coppicing rotations in order to obtain about 10-12 Mg ha-1 year-1 of dry above-ground biomass ([3], [28], [5], [12], [10], [39], [13]). Several experiments have obtained new or already-in-use clones for biomass production ([21], [4], [36]). Hence, the biennial and annual yield of SRF has been extensively assessed by several studies ([22], [35], [25]), many of which have focused on clonal poplar experiments, aimed at testing the productivity and pests resilience ([14]) under contrasting latitudinal sites and experimental trials ([18], [20], [26]), but few studies have continuously monitored the growth of a SRF during its vegetative cycle.
The general objective of this work is the assessment of productive characteristics of three selected poplar clones (AF2, AF6 and Monviso) to site conditions of central Italy, as affected by the rotation cycles, as well as by the planting density, comparing low- (7 140 cuttings ha-1) vs. high-density (10 360 cuttings ha-1) plantations.
Materials and methods
Site descriptions, plant material and experiment layout
The experimental plantation (total surface: 4 ha) was established in 2005 within the farm of the Agricultural Engineering Research Unit of the Agricultural Research Council (CRA-ING), 30 km east of Rome (Monterotondo; 42° 06’ N, 12° 37’ E), on a flat field, with a clay-loam soil texture and a low level of soil organic matter, nitrogen and phosphorus ([8]). Both pre- and post-planting agronomic practices are shown in Tab. 1, according to the recommended management practices in Italy ([11]). The plantation was only irrigated with an emergency-irrigation in July 2005 (1400 m3) to promote a better rooting of cuttings. Cuttings had a length between 220 and 280 mm with a maximum diameter of 28 mm. In order to favor the coppice’s growth, a “cut back” at the end of the first vegetative season (2005) was carried out. At this date the average yield of the plantation was 1.9 MgDM ha-1.
Tab. 1 - Main agronomic practices in the studied plantation.
| Proceding crop | Wheat |
| Plowing deep (cm) | 80 |
| Fertilization (kg ha-1) | 100 P2O+100 K2O |
| Harrowing deep (cm) | 15 |
| Planting date | March 2005 |
| Chemical weed control | Post-emergence |
| Top dressing (kg ha-1) | 100 N |
| Irrigation (m3 ha-1) | 350 (July 2005) |
| Cut-back date | February 2006 |
| Harvesting date | December 2007; March 2011 |
Meteorological data (average annual temperature and annual precipitation) were collected from a nearby station, located few meters from the plantation. These data were calculated over the five observed years (from 2006 to 2010) and compared with the last ten years (from 2001 to 2010 - Tab. 2). Annual precipitation reached a maximum of 927.2 mm for the year 2009 and a minimum of 473.0 mm for the year 2007. May-August precipitation values were similar among years (except 2007), with a mean value of the last 10 years of 42.3 mm. Average annual temperature during the study period was 15.7 °C, whereas for the same period the average monthly temperature for July was 25.4 °C, with a mean value of the last 10 years of 15.3 °C. Drought period was shorter in the years 2008-2010 (3.5 months) than in years 2006-2009 (5 months).
Tab. 2 - Summary of main meteorological characteristics during the study period (2006-2010) and for the last ten years (2001-2010). Data and metadata of PET (potential evapo-transpiration for Monterotondo site) were extrapolated from the database of the Research Unit for Climatology and Meteorology applied to Agriculture (CRA-CMA). Source: ⇒ http://old.politicheagricole.it/ucea/forniture/index3.htm
| Year | Annual Rainfall (mm) |
Rainfall May-August (mm) |
PET (mm) |
Aridity Index (P/PET) |
Average Annual Temp (°C) |
Average Temp July (°C) |
|---|---|---|---|---|---|---|
| 2006 | 512.6 | 30.3 | - | - | 15.8 | 27.2 |
| 2007 | 473 | 18.5 | - | - | 16.5 | 25 |
| 2008 | 780.5 | 49.5 | - | - | 15.5 | 24.5 |
| 2009 | 927.2 | 65.4 | - | - | 15.6 | 24.5 |
| 2010 | 905.3 | 75.4 | - | - | 15 | 25.7 |
| 2001-2010 | 627.5 | 42.3 | 1010 | 0.62 | 15.3 | 25.4 |
Three poplar clones were evaluated: P. x canadensis Moench “AF2”, P. nigra L. x P. x generosa A. Henry “AF6”, P. x generosa A. Henry x P. nigra L. “Monviso” ([27]). They are the most trade between those selected for biomass production and for their resistance to natural infections of Melampsora spp. and Marssonina brunnea. To test the influence on growth of density/ spacing, the plantation was established with two different planting designs, differing in densities and spacings, both as single and twin rows, hereafter called “SR” and “TR”, respectively; (i) low density (7 140 cuttings ha-1) were placed in a SR (2.80 m between the rows and 0.50 m along the row); (ii) high density (10 360 cuttings ha-1) were placed in a TR (2.80 m between twin-rows, 0.75 m between the row forming a pair, 0.50 m along the rows).
The experimental data were recorded from 2007 to 2010, in order to test two experimental treatments (factors): (i) two planting densities/spacings (main plot); and (ii) three clones (sub-plot). Each clone of SR was replicated six times on elemental plots (18 plots totally) with a unit surface area of 67.2 m2. In the TR, each clone was replicated four times (12 plots totally) with a unit surface area of 57 m2. The size of SR and TR plots were respectively 8.4 m x 8 m (three rows and 16 stumps per row) and 7.1 m x 8 m (two twin-rows and 32 stumps per row).
Field performance measurements (growth and yield)
Field performance was evaluated yearly by measuring the diameter at breast height (DBH) of all shoots in each replicated plot. Total height (H) was measured on thirty shoots for each clone, in each replicated plot. A tree caliper (Silvanus type 1208, accuracy 0.5 cm) was used for measuring the DBH and a logger’s tape for determining tree height after felling.
In order to assess the yields for each clone, allometric relationships between fresh weight and DBH were determined in both SR and TR using non-linear regressions according to Al Afas et al. ([1]) and Verani & Sperandio ([39]) as follows (eqn. 1):
where Y is the fresh weight of shoot (kg), x is the DBH (cm) and a and z are the regression parameter to be determined. To estimate the parameters a and z, a destructive sampling of 180 living trees (30 shoots x 3 clones x 2 density) was made. To obtain the dry weight from fresh weight, the moisture content on 45 shoots (15 per clone) was determined. Shoot fresh weight was measured on woody disks taken from basal, middle and upper parts of the stem just before each coppicing cycle. Woody disks were immediately weighed in the field with a precision scale (Orma model BC16D) and then taken into the laboratory for moisture determinations, according to the thermo-gravimetric methods ([38], [37]). The results of non-linear regressions between fresh weight and DBH are reported in Tab. 3. Except the clone AF6 of TR in 2007 (R3S2), observed coefficients of determination (R2) from regression analysis were always larger than 0.8. Yield (expressed in MgDM ha-1) of each clone was obtained by multiplying the estimated weight (using the specific allometric equation) of each single shoot for the number of the shoots per hectare.
Tab. 3 - Relationships between fresh weight and DBH for each clone and planting design observed at the end of the two coppicing rotations.
| Planting Design |
Clone | 2007 (R3S2) | 2010 (R6S3) | ||||
|---|---|---|---|---|---|---|---|
| a | z | R2 | a | z | R2 | ||
| SR - Low density |
AF2 | 0.252 | 1.94 | 0.84 | 0.363 | 1.92 | 0.93 |
| AF6 | 0.17 | 2.25 | 0.84 | 0.302 | 2.09 | 0.96 | |
| Monviso | 0.136 | 2.52 | 0.96 | 0.327 | 1.98 | 0.92 | |
| TR - High density |
AF2 | 0.218 | 2.01 | 0.92 | 0.451 | 1.69 | 0.94 |
| AF6 | 0.188 | 2.19 | 0.63 | 0.488 | 1.81 | 0.94 | |
| Monviso | 0.335 | 1.84 | 0.9 | 0.939 | 1.21 | 0.92 | |
Field performance in the following growing stages was evaluated as: roots aged 3 years and stems aged 2 years (R3S2, year 2007 just before the first harvesting); roots aged 4 years and stem aged 1 year (R4S1, vegetative season 2008); roots aged 5 years and stems aged 2 years (R5S2, vegetative season 2009); roots aged 6 years and stems aged 3 years (R6S3, vegetative season 2010 just before the second harvesting).
Statistical analysis
Data collected on DBH, H, yield, number of shoots ha-1 and shoots/stump ratio were compared among clones and analyzed by two-way ANOVA in each separate year using PRISM software version 4.01 (GraphPad Software, Inc. San Diego, CA, USA). Measurements on individual trees were used as observations, whereas densities (2 levels) and clones (3 levels) were defined as factors. When significant differences (α = 0.05) between means were detected, post-hoc tests were performed using Tukey HSD test for determining significance. Pairwise Student’s t-test was used to test differences in wood moisture content between R3S2 and R6S3.
Results
Effect of clone on growth and yield
Variability among clones was observed in most of the studied parameters in R3S2, R4S1, R5S2 and R6S3 respectively (Tab. 4 and Tab. 5). Apart R3S2 where non-significant differences in DBH and stool survival were detected, AF2 clone usually has always shown a better growth performance compared to AF6 and Monviso clones. Yield interactions between density and clone were significant in both R3S2 and R6S3 and reflected the much greater increases for TR in R3S2 and for clone AF2 in R6S3.
Tab. 4 - Main effect of experimental treatment on both growth and yield at the end of the biennial rotation (R3S2). Means in vertical sequence not followed by the same letter are significantly different at P = 0.05, according to Tukey’s HSD test. (ns): not significant; (*): P = 0.05; (**): P = 0.01; (***): P = 0.001.
| Treatment | Growth year 2007 (R3S2) | |||||
|---|---|---|---|---|---|---|
| DBH cm |
Height m |
Shoots plant ha-1 |
Shoots/ Stump |
Survival % |
Yield Mg ha-1 y-1 |
|
| AF2 | 2.6 a | 4.8 a | 28 552 a | 3.6 a | 92.2 a | 11 a |
| AF6 | 2.7 a | 4.6 a | 28 197 a | 3.8 a | 90.6 a | 10.8 a |
| Monviso | 2.7 a | 5.3 b | 21 780 b | 2.9 a | 92.5 a | 8.8 b |
| TR - High density | 2.9 | 2.7 | 24 214 | 2.6 | 89.3 | 11.8 |
| SR - Low density | 2.4 | 4.8 | 28 180 | 4.2 | 94.2 | 8.5 |
| Average | 2.7 | 4.9 | 26 176 | 3.4 | 91.7 | 10.1 |
| Clone | ns | *** | ** | ns | ns | * |
| Density | ns | ns | ** | ** | * | ** |
| Clone x Density | ns | ** | ** | ns | ns | *** |
Tab. 5 - Main effect of experimental treatment on both growth and yield at the end of the triennial rotation (R6S3). Means in vertical sequence not followed by the same letter are significantly different at P = 0.05 according to Tukey’s HSD test. (ns): not significant; (*): P = 0.05; (**): P = 0.01; (***): P = 0.001.
| Treatment | 2008 | 2009 | 2010 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1st year (R4S1) | 2nd year (R5S2) | 3th year (R6S3) | ||||||||
| DBH cm |
Height m |
DBH cm |
Height m |
DBH cm |
Height m |
Shoot plant ha-1 |
Shoot/ Stump |
Survival % |
Yield Mg ha-1 y-1 |
|
| AF2 | 1.8 | 3.7 | 2.6 | 6.7 | 3.3 | 6.7 | 27 551 | 3.3 | 90.2 | 13.9 |
| AF6 | 1.4 | 3.2 | 2.2 | 6.0 | 3.1 | 5.7 | 25 612 c | 3.4 | 84.3 | 13.2 |
| Monviso | 1.5 | 3.3 | 2.0 | 6.4 | 2.8 | 6.7 | 26 639 | 3.5 | 88.9 | 12.0 |
| TR- High density | 1.5 | 3.4 | 1.9 | 5.7 | 2.7 | 6.2 | 28 840 | 3.0 | 85.6 | 12.2 |
| SR - Low density | 1.6 | 3.4 | 2.3 | 6.3 | 3.4 | 6.6 | 24 361 | 3.8 | 89.9 | 13.9 |
| Average | 1.5 | 3.4 | 2.1 | 6.0 | 3.1 | 6.4 | 26 601 | 3.4 | 87.8 | 13.0 |
| Clone | *** | *** | *** | *** | ** | ** | * | ns | * | * |
| Density | ns | ns | ** | *** | ** | ns | ** | ns | * | * |
| Clone x Density | ** | ns | ns | ** | ns | ** | ** | ns | ** | ** |
Considering that the moisture content observed in R3S2 and R6S3 showed non-significant differences among clones and among densities with an average value of 54.2% (Tab. 6), yields obtained from both R3S2 and R6S3 showed significant differences among clones (P<0.05 for both). This was because AF2 clone gave the highest mean yield (11.0 MgDM ha-1 year-1), whereas Monviso the lowest mean yield (8.8 MgDM ha-1 year-1) in R3S2. Similarly, yield of AF2 clone also reached highest mean values in R6S3 (13.9 MgDM ha-1 year-1) compared to both AF6 and Monviso clones (13.2 and 12.0 MgDM ha-1 year-1 respectively). This was because at the end of the triennial rotation, AF2 showed both highest stump survival (90.2%) compared to AF6 (84.3%) and Monviso (88.8%), and highest number of shoots (27 751 trees ha-1) compared to those of the other two clones (25 612 and 26 183 shoots ha-1 for both AF6 and Monviso).
Tab. 6 - Moisture content observed in R3S2 and R6S3.
| Moisture content % | 2007 (R3S2) |
2010 (R6S3) |
|---|---|---|
| AF2 | 52.41 | 55.96 |
| AF6 | 53.01 | 55.62 |
| Monviso | 53.63 | 54.62 |
Effect of density on growth and yield
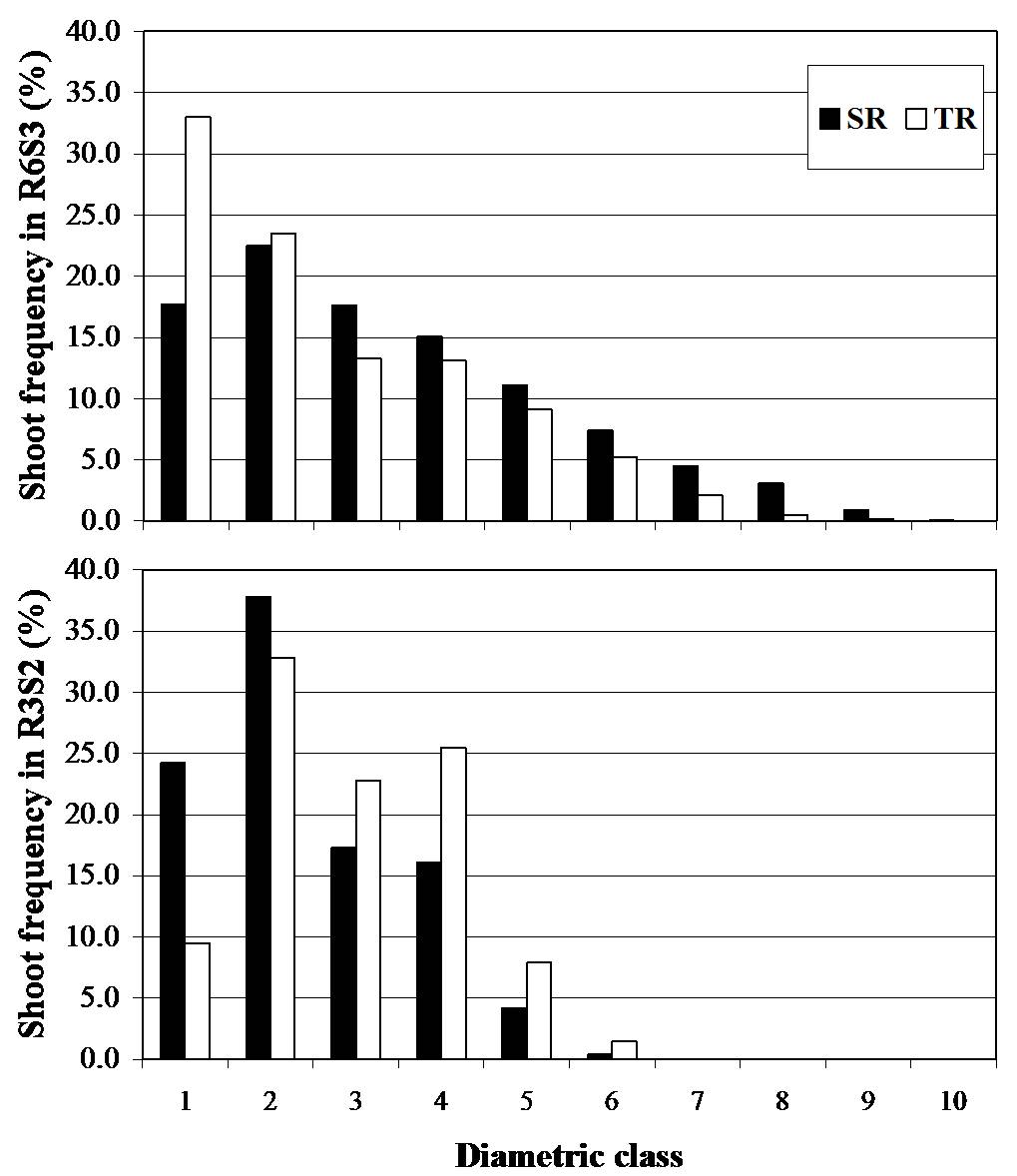
Density had significant effects on both growth and yield in R3S2, R5S2 and R6S3, whereas non-significant differences were found in R4S1. In R3S2 we observed a density effect on shoots/stump ratio with highest values observed in SR compared to TR. At the end of the triennial rotation (R6S3) the SR showed higher yield (13.9 MgDM ha-1 year-1) compared to TR (12.2 MgDM ha-1 year-1). Conversely, at the end of the biennial rotation (R3S2) the highest yield was obtained in TR (11.8 MgDM ha-1 year-1) compared to SR (8.5 MgDM ha-1 year-1). Density effect on growth (DBH and H) was time dependent as indicated by two different growth phases. The first phase was characterized by the absence of a density effect, which lasted 2 years (i.e., R3S2 and R4S1). Conversely, the second phase was characterized by an increase in R5S2 and R6S3. This could be explained by the fact that TR showed lowest values in growth, due to the intra-specific competition more prolonged in R5S2 and R6S3 than in R4S1. DBH increased predictably as density between trees decreased, as already reported ([2]), especially for the triennial rotation. Interestingly, total height (H) increases as density decreased for both coppicing rotations. Variations in height growth with changes in available growing space could be attributed to ontogeny, to the range of tested spacing treatments, or to species. This was because height growth plays an important role in morphological acclimation to light competition ([19]), with plants tending to allocate more photosynthate to height than diameter growth, which results in increasing stem slenderness. Moreover we observed a change in distribution among diametric classes of the shoot populations according to the two harvesting date at the end of the first (R3S2) and second coppicing rotation (R6S3). This was because there was a shift towards higher frequencies in the lower diametric classes in R6S3 (Fig. 1), such as for shoots of the first two diametric classes of TR (Fig. 2), with negative consequences on qualitative characteristics of the woody biomass, likely due to an higher bark percentage ([16]). This response could explain the occurrence of larger height growth in SR, and faster shifts vs. higher diametric classes in R6S3 and TR, highlighting a different trade-off between mechanical stability of the stem and height growth.
Fig. 1 - Shoot DBH distribution among clones observed at the end of the two coppicing rotations (i.e., R3S2 and R6S3).
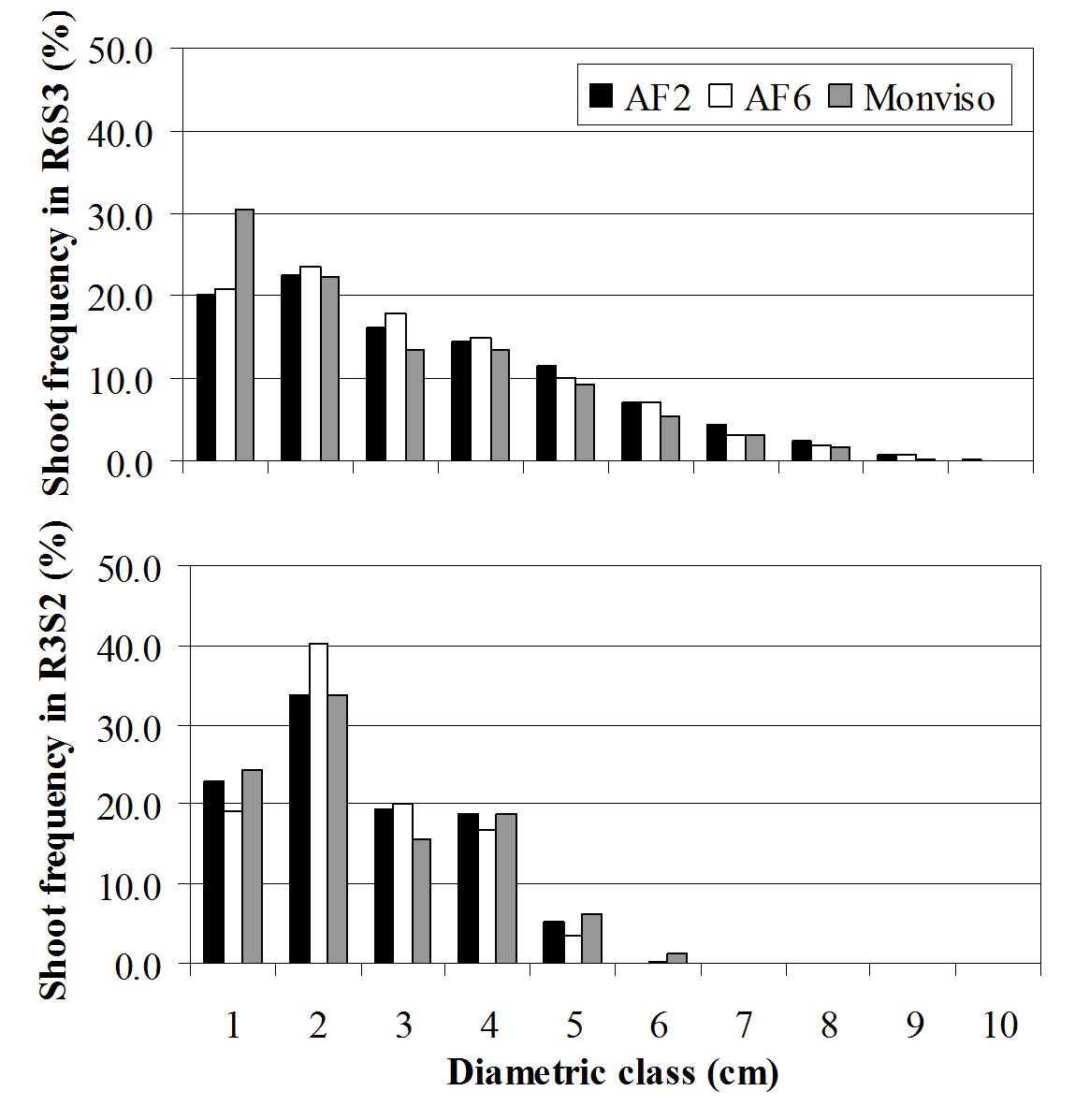
Fig. 2 - Shoot DBH distribution among planting designs observed at the end of the two coppicing rotations (i.e., R3S2 and R6S3)
Discussion
Poplar’s cultivation for yield production in central and southern Italy is limited by spring and summer drought. Of course, even in optimal climatic locations, the SRF cultivation should be always accompanied by cultural operations, such as irrigation and fertilization, for improving soil fertility. Optimal conditions for hybrid poplars cultivation in Italy are: (i) mean annual temperature: 8.5 to 17 °C; (ii) summer precipitation: 100-150 mm. In our case-study, local climatic conditions were outside the optimum for poplars (aridity index of 0.62 mm). Indeed, we observed very long summer drought (average 2006-2010: up to 4.0 months), and May-August precipitation always below 80 mm, with an average value of the last ten years of 40.2 mm. Therefore, the case-study is a sub optimal site for cultivation of poplar species. Yearly above-ground yields in SRF plantations for the same hybrid poplar clones planted in northern Italy (along the Po valley) are greater, ranging between 5.6 and 17.9 MgDM ha-1 year-1 ([26]). Nevertheless, the yearly yield obtained in the plantations in northern Italy are similar to those observed in boreal regions (central Europe, USA, and Canada) not affected by summer dryness ([3], [9]). Contrastingly, lower yearly productions were observed by Minotta & Muzzi ([22]) and Pannacci et al. ([25]) in SRF plantations of central Italy, with yields ranging between 1.3 and 6.2 MgDM ha-1 year-1. The results obtained in this study are surprisingly similar to those reported for boreal (cold temperate) conditions, ranging between 10.1 (at the end of the first coppicing rotation) and 13.0 MgDM ha-1 year-1 (at the end of the second coppicing rotation). Christersson ([7]) reported for Sweden an average yearly yields of 3-10 MgDM ha-1 year-1 in 5-year-old plantations, while Willebrand et al. ([42]) reported average yearly yields varying between 8 and 14 MgDM ha-1 year-1 in 6-year-old plantations. In spite of the short duration of the growing season and low soil nitrogen in boreal regions, a yield of 20 MgDM ha-1 year-1 could easily be achieved in our plantation by adding fertilizers and selecting appropriate clones and spacings, as reported for Swedish plantations by Weih ([41]). This in fact would explain the high potential of the studied poplar clones, even in Mediterranean sites affected by summer drought.
Conclusion
In southern Europe, the use of SRF plantations appears to offer a highly promising and politically desirable option among the range of renewable energy sources currently available. SRF has been adopted mostly in northern Italy owing of the favorable climatic and soil conditions for poplars cultivation. Demand for woody biomass for fuel, energy purposes or fiber combined with reduced land availability has forced land managers to increase productivity of SRF plantations. Under such circumstances new high-yielding poplar clones were tested for growing in areas of central Italy affected by Mediterranean climatic conditions. Our study has shown that the climate of central Italy offers favorable conditions for the biomass production of selected hybrid poplar clones, highlighting very high productive potential when summer dry periods are frequent (especially from May to August) and temperatures are less mild (slightly higher than the normal).
The poplar clones tested had the best performances in terms of growth and yield when planted in a SR design during the second coppicing rotation. Therefore SR (7 140 cuttings ha-1) was more advisable than TR (10 360 cuttings ha-1) in order to obtain good biomass production with low planting costs. At clone-level, AF2 showed higher DBH values and yield productions than AF6 and Monviso in both biennial (R3S2) and triennial (R6S3) rotations.
Significant functional relationships between fresh biomass production and DBH, appropriate for this climatic area and environmental conditions, for every poplar clone were found. These may be used for a non-destructive prediction of dry yield and for rapid yield estimations of the shoot populations.
Acknowledgments
Funds for this study were provided by the Italian Ministry of Agriculture Food and Forest Politics (D.M. 487/7303/2004) within the COFEA project (Costituzione di una Filiera Energetica di Autoconsumo) and developed within the post-doctoral research activities of Giovanni Di Matteo on biomass productions for energy purposes in Short Rotation Forestry of central Italy.
References
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
S Verani
Consiglio per la Ricerca e la Sperimentazione in Agricoltura, Unità di ricerca per le Produzioni Legnose fuori Foresta (CRA-PLF), v. Valle della Quistione 27, I-00166 Rome (Italy)
Consiglio per la Ricerca e la Sperimentazione in Agricoltura, Unità di ricerca per l’Ingegneria Agraria (CRA-ING), v. della Pascolare 16, I-00015 Monterotondo, RM (Italy)
Corresponding author
Paper Info
Citation
Di Matteo G, Sperandio G, Verani S (2012). Field performance of poplar for bioenergy in southern Europe after two coppicing rotations: effects of clone and planting density. iForest 5: 224-229. - doi: 10.3832/ifor0628-005
Academic Editor
Renzo Motta
Paper history
Received: Jan 24, 2012
Accepted: Aug 20, 2012
First online: Oct 02, 2012
Publication Date: Oct 30, 2012
Publication Time: 1.43 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2012
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 58546
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 48653
Abstract Page Views: 3471
PDF Downloads: 4855
Citation/Reference Downloads: 28
XML Downloads: 1539
Web Metrics
Days since publication: 4784
Overall contacts: 58546
Avg. contacts per week: 85.67
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2012): 35
Average cites per year: 2.50
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Biomass production of Populus nigra L. clones grown in short rotation coppice systems in three different environments over four rotations
vol. 7, pp. 233-239 (online: 10 March 2014)
Research Articles
Preliminary study on genetic variation of growth traits and wood properties and superior clones selection of Populus ussuriensis Kom.
vol. 12, pp. 459-466 (online: 29 September 2019)
Research Articles
Climate-wise models of biomass productivity for hybrid poplar clones in Europe
vol. 16, pp. 188-194 (online: 30 June 2023)
Research Articles
Nursery screening of poplar and willow clones for biofuel application in Ukraine
vol. 15, pp. 401-410 (online: 06 October 2022)
Research Articles
Interaction between planting spacing and wood properties of Eucalyptus clones grown in short rotation
vol. 14, pp. 12-17 (online: 02 January 2021)
Research Articles
Variation of wood and bark density and production in coppiced Eucalyptus globulus trees in a second rotation
vol. 9, pp. 270-275 (online: 08 September 2015)
Research Articles
First vs. second rotation of a poplar short rotation coppice: leaf area development, light interception and radiation use efficiency
vol. 8, pp. 565-573 (online: 27 April 2015)
Research Articles
Nitrogen removal and its determinants in hybrid Populus clones for bioenergy plantations after two biennial rotations in two temperate sites in northern Italy
vol. 8, pp. 668-676 (online: 02 February 2015)
Research Articles
Effective woody biomass estimation in poplar short-rotation coppices - Populus nigra × P. maximowiczii
vol. 16, pp. 202-209 (online: 25 July 2023)
Short Communications
Effect of intensive planting density on tree growth, wood density and fiber properties of maple (Acer velutinum Boiss.)
vol. 9, pp. 325-329 (online: 22 October 2015)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







