Genetic monitoring in forests - early warning and controlling system for ecosystemic changes
iForest - Biogeosciences and Forestry, Volume 4, Issue 2, Pages 77-81 (2011)
doi: https://doi.org/10.3832/ifor0571-004
Published: Apr 05, 2011 - Copyright © 2011 SISEF
Research Articles
Collection/Special Issue: IUFRO RG 7.01 2010 - Antalya (Turkey)
Adaptation of Forest Ecosystems to Air Pollution and Climate Change
Guest Editors: Elena Paoletti, Yusuf Serengil
Abstract
In the context of existent programmes of environmental monitoring which have been established as efficient tools for permanently observing environmental changes, a concept for a genetic monitoring in forests was recently elaborated by a German forest geneticist working group. Genetic monitoring is assumed to contribute essentially to the estimation and valuation of the effect of factors influencing the genetic system of trees in the forests, thus making it an early warning and controlling system for ecosystemic changes. The “Concept of a Genetic Monitoring for Forest Tree Species in the Federal Republic of Germany” gives scientifically-based guidelines for monitoring the current state and dynamics of genetic systems in forest stands of diverse tree species in an extensively, harmonized manner. Both objectives and the realization of the genetic monitoring concept are presented here. The status of the genetic systems of forest tree populations is assessed on the basis of criteria, indicators and verifiers. For this purpose the genetic as well as the phenological and physiological levels are taken into consideration in order to follow temporal developments and to estimate influencing factors. The results of a pioneer study concerning the tree species Fagus sylvatica and Prunus avium are reported.
Keywords
Genetic monitoring, Fagus sylvatica, Prunus avium, Indicators, Mating system
Introduction
The conservation of genetic resources is an important issue in managed forests. The adaptability of trees to environmental factors such as climate change or to outbreaks of pests and diseases depends basically on genetic variation. Thus, it is crucial for the maintenance of forests to prevent the erosion of genetic diversity.
Genetic diversity with its inherent extent of allelic and genotypic structures is the elementary base for biological diversity. It is considered to be one of the criteria which are most important for assessing the potential for dramatic and potentially deleterious changes for any forest tree stand during its lifetime. Especially under the present impact of predicted climate change, it is necessary that forest ecosystems with high levels of biodiversity are capable of persisting and surviving. Even more it is an essential prerequisite to safeguard the diverse functions of the forests in a future-oriented way. In this context it is of top priority to maintain genetic diversity and to protect the genetic system of the woody plant species.
The recent increases in temperature resulting from global climate change impose a directional selection pressure on the population, which generates a change of the genotype frequencies. An evolutionary response to warming temperatures can be expected ([12]). As a consequence, many efforts are underway presently on the international and national levels to conserve forest genetic resources both in situ and ex situ in a sustainable way. The manifold collaborating activities among different international organizations, programmes and processes like IUFRO, EUFORGEN, the CBD-process, IPGRI, and some more should be stressed in this context.
Great efforts are practiced also on the level of individual nations. In Germany the “Concept for the Conservation of Forest Genetic Resources” was already introduced in 1987 and was revised thoroughly in 2000 (see ⇒ http://www.genres.de/fgrdeu/blag). The latter version by Paul et al. ([18]) includes expressis verbis for the very first time the target to develop of a long-term genetic monitoring scheme. The long-term potential of survival for genetic reserves should also be assessed ([5]).
While some diverse environmental monitoring programmes have been established in the past 10 to 15 years as efficient tools for permanently observing environmental changes, genetic aspects are lacking however. Information from monitoring programs could be used to update risk assessments in forest management plans in an adaptive-management sense. The task for the next decade is to understand better how climate affects tree population and how forests respond to them. Improved monitoring programs and analytic tools are needed to develop this understanding ([6], [22]).
As a result of this situation a concept for a genetic monitoring scheme in forests was elaborated by a German forest geneticist working group (see [3], [4]; [14], [15]). This decisive measure is regarded as an essential step towards implementing the Convention on Biological Diversity which in paragraph 7b calls for the surveillance of the distinct constituents of biological diversity ([1]).
To the best of our knowledge, it is the very first time that a concept for a genetic monitoring in forests was elaborated and tested in pilot studies. Similar considerations exist only for different plant and animal species ([21]). However, we do not know whether or not such approaches have been realized in the meantime.
Objectives of a genetic monitoring scheme
The aim of carrying out a genetic monitoring is to observe the dynamics of transition from the present to the future genetic status of a forest stand. In order to do this, the status of the genetic system must be conceived in its entirety. The genetic system comprises all mechanisms and processes which contribute to the production, the maintenance, and last but not least to the modification of genetic variability and its transfer to the succeeding generations. Initially the essential attributes (characteristics) of this system must be detected. Then parameters or indicators are needed which clearly show whether this system is intact or disturbed, and to what extent. Finally the specific hypotheses are examined using verifiers.
The German concept for genetic monitoring of forest tree species
Indicators and verifiers
The intactness (resp. functionality) of the genetic system of a species is characterized by the population genetic processes of selection, mating system and migration. Thus, the maintenance of these processes represents the criteria for conserving genetic variation. The German “Concept for Genetic Monitoring in Forests“ as published in 2004 by the BLAG-Expertengruppe “Genetisches Monitoring” is based on the indicators proposed by Namkoong et al. ([17]) for population genetic processes concerning the following points:
- level of genetic variation;
- directional change in gene or genotypic frequencies;
- changes in mating system processes;
- gene migration between populations.
Genetic monitoring uses these indicators to assess the conditions and the dynamics of the processes of the genetic system by repeating this procedure at different times ([11]).
For this procedure, common population genetic data and the parameters deduced from this data are estimated. These data represent the verifiers of the given indicators. Tab. 1 summarizes keywords characterizing the verifiers for the respective indicators.
Tab. 1 - Indicators and verifiers of the genetic processes. (P): number of polymorphic loci; (A/L): mean number of alleles per locus; (F-value): fixation index.
| Indicators | Verificators |
|---|---|
| Level of genetic variation | Gene frequencies, genetic diversity, P, A/L, variation in phenological parameters, F-value,Number of potential parent trees*) |
| Directional change in gene or genotypic frequencies | Differences between cohorts: - in allele, genotype and phenotype frequencies, - in the distribution of age classes |
| Changes in mating system processes | Mating system, rate of cross-fertilization, rate of biparental inbreeding, number of effective pollen donors, proportion of empty/full seed and germinability |
| Gene migration between populations | Dispersion of pollen and seeds, differentiation between populations of the same age, isolation, family structures |
Gene frequencies and different genetic parameters based on gene frequencies, like genetic diversity, number of polymorphic loci, mean number of alleles per locus, variation in bud burst, in frost resistance etc. can be applied efficiently as measures for assessing the level of genetic variation. The outcrossing rate, the number of effective pollen donors, but also the quality of seed reflected by the proportion of empty seed or by germinability supply valuable information about the mating system. Gene migration is verified by the pollen and seed dispersal distance as well as by the presence/absence of family structures within a tree population.
The different verifiers that can be measured and are used for any indicator differ widely in their individual precision by which they estimate the genetic parameters. And last but not least, they differ in cost and the numbers of facilities required to sample and measure them.
Realization
Genetic monitoring includes the periodical, randomized or plot-specific inspection of genetic structures and characteristics of the genetic system (e.g., population size, flowering and sexual system). It should be carried out both in managed and unmanaged forests.
The following strategy for carrying out genetic monitoring in practice is proposed in the concept:
- identifying the genotypes of the reproductive trees in the selected monitoring plots or stands through a basic inventory using genetic markers;
- assessing the genetic composition of natural regeneration of an adequate sample from the stand under study; this procedure should be repeated after 10-15 years;
- performing additional inventories periodically to determine the extent of flowering and fructification of selected trees (e.g., every 1-2 years);
- carrying out additional genetic studies on the seeds from different seed years (e.g., every 5 years);
- calculating the given indicators for genetic processes based on the genotypes of the adult trees, natural regeneration and the seeds;
- simulating, the temporal and spatial dynamics of the genetic and demographical composition of tree populations based on empirical monitoring data in order to identify any critical development,
The tree species are chosen according to their importance concerning usability, existence, and indicator capacity. It is recommended to work first on tree species for which genetic markers are available for routine serial tests. For reasons of comparability, genetic monitoring should be performed according to tree species-specific harmonized methods, particularly regarding sampling strategy, inventory procedures, analyzing techniques, documentation, and data evaluation. The data obtained is to be recorded in such a way that it will be useful in simulation processes.
Both managed and unmanaged sites may be used as monitoring plots. A detailed documentation must be made to ensure continuity of the monitoring procedure. The geographic coordinates of the plot, the history of the stands and the treatments carried out should be considered. Furthermore, the geographic coordinates of the individual trees, their quality traits as well as their social classification must be considered. If possible, relevant environmental data like weather, atmospheric pollution, quantity of pollutant input, and soil vegetation should be recorded.
Pilot studies
Methodological aspects
In Germany the first practical experiences were gained in a pilot study conducted on European beech (Fagus sylvatica) as an important predominant wind-pollinated species ([16]) and wild cherry (Prunus avium) as a rare species pollinated by insects ([9]). This study included four beech stands located in north (N), east (E), central (C), and south Germany (S) and five wild cherry populations. The wild cherry stands under study include the stands Einhaus (northern Germany); Chorin (north-eastern Germany); Spargründe (eastern Germany); Hildesheimer Wald (central Germany), and Reutlingen (southern Germany).
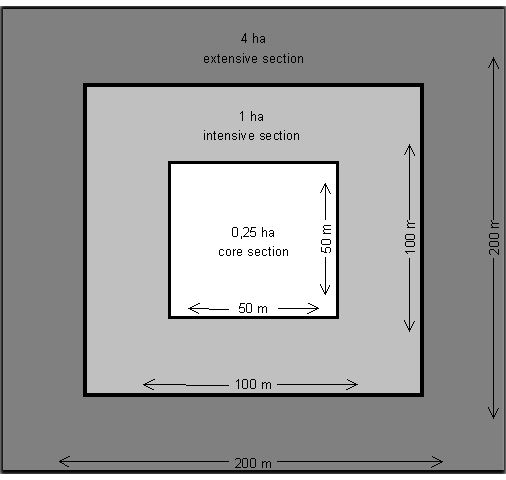
Beech - Each experimental beech plot includes a total area of 4 hectares, subdivided into 3 zoned sections which are marked visibly by pegs. There is a central part of 50 x 50 meters (“core section”, 0.25 ha), surrounded by a square of 100 x 100 meters which is called the “intensive section”.
It comprises a total area of 1 ha. Within the core zone particularly additional parameters like flowering phenology, leaf unfolding and nutrient analyses were monitored. Outside of this intensive plot, an area of 200 x 200 meters is delimited which is called the “extensive section”. The overall area is 4 ha (Fig. 1). Sampling of adult trees, natural regeneration and seeds was performed within these different zones at differing intensities.
Fig. 1 - Schematic design of a monitoring plot for abundant tree species presenting the arrangement in core, intensive and extensive sections.
The collected materials were used for genetic studies and for characterizing the quality structure of beechnut seeds (see Tab. 2). The position of each sample tree was determined exactly by applying for example GPS.
Tab. 2 - Overview of the sampling procedure to be carried out in the 3 sections of a monitoring plot.
| Developmental state | Intensive section including core section | Extensive section |
|---|---|---|
| Genetic Studies | ||
| adult trees | all individuals present | additional individuals up to max. 300 |
| natural regeneration | 200 young plants representative for the overall plot | |
| 4 clusters of natural regeneration comprising 50 individuals each | - | |
| beechnuts seeds | collecting per individual tree separately beechnuts from 20 adult trees | |
| Quality structure of beechnut seeds | ||
| beechnuts seeds | seed mixture of the overall stand | |
For genetic studies isozyme gene markers as well as molecular genetic markers including microsatellites (SSRs) and Arbitrary Fragment Length Polymorphisms (AFLPs) were applied. Different genetic parameters were calculated reflecting genetic variation (verifiers - see Tab. 1), spatial genetic structures, parameters reflecting the mating system, pollen distribution functions, and parental analysis. Programs used included GDA-NT ([8]), GSED ([10]), SGS-Spatial Genetic Software ([7]), MLTR-Multilocus Mating System Program ([19]), PolDisp ([20]), and Cervus ([13]).
Random samples of the beechnut seeds collected within the four monitoring plots were tested for their quality structure according to the ISTA regulations ([2]). Phenological observations included basically flushing in springtime which was carried out on selected individual trees.
Prunus - Because of its low and variable density, the plot size for Prunus avium was defined as the area that includes 150 reproductive trees. The size of the five wild cherry plots varied from 0.6 ha to 18.1 ha depending on the variable densities of reproductive trees (5 individuals per ha to 230 individuals per ha). In each plot 150 saplings were sampled in a statistically representative way over the whole area. Seeds were collected from at least 30 trees within each plot. The genetic inventories were done by applying nuclear microsatellites, AFLPs and markers for the known locus of the self-incompatibility (S-locus). Furthermore the spatial position, diameter, natural social class, and flowering phenology for of the reproducing trees were measured and the quality of seeds was tested. For computing the genetic parameters the same analysis programs were used as for beech.
First results
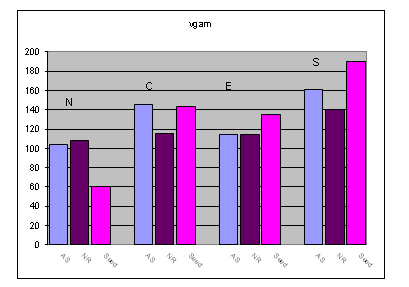
Beech - Except for plot N the differences in genetic diversity between the three developmental stages of the stands under study were found to be low (see Fig. 2).
Fig. 2 - Genetic diversity (expressed as vgam) as determined in the four beech monitoring plots for the adult stand (AS), natural regeneration (NR) and the seeds.
The outcrossing rate exceeded 95%, the number of effective pollen donors was found to be high, at 12 to 63 trees. The maximum pollen dispersal distance varied from 112 to 133 m. Consequently, the mating system appears to be widely intact. Moreover, a high potential for transmitting genetic information over generations can be assumed. Repeated inventories to be performed in the following years will reveal whether or not this intactness will be sustained.
The four monitoring plots comprise different distinct gene pools which represent each local gene pool well. This observation makes it clear that plots located in different regions must be included when performing a genetic monitoring in order to consider local peculiarities,
Differences in flushing became evident both in the years of observation and in all of the four monitoring plots. In 2006 flushing started earlier in plot S as compared to plot E, and this process took about six more days. In 2007 flushing started on both plots significantly earlier. These observations will be pursued in the next years in order to make specific evaluations associated with plot-related climate data.
The combination of different indicators and verifiers was used as an approach to assess the intactness of the genetic system. A set of 18 indicators/verifiers was applied according to the Concept of Genetic Monitoring. It included the:
- level of genetic diversity in the mature stand verified by the effective numbers of alleles of the analyzed isozymes, SSRs and AFLPs as well as the level of heterozygosity of the isozymes, and flushing;
- changes in the gene and genotype frequencies verified by estimating the differences between the developmental stages of the trees within a stand and the differences between the cohorts of natural regeneration;
- mating system verified by a deficit of heterozygous mature individuals as compared to Hardy Weinberg equilibrium (isozymes); open pollination rate; number of effective pollen donors; spatial genetic structures (family structures); and germination percent of the seeds in 2006; and
- migration verified by the differences between the genetic differentiation of the mature stands studied and its pollen dissemination in 2006.
Each verifier was assigned one of the three values, either -1, zero (0), or +1. +1 signifies an expected positive effect on the genetic system of the tree population. -1 indicates a negative effect and 0 was assigned in case of incomplete data. The average of all verifiers was used to judge the overall situation of each population. Based on this, the genetic system present now was appraised undisturbed for two out of the four monitoring plots (plot N and plot S). No clear statement could be made for plot E. Therefore clarity must be obtained by carrying out another survey. For plot C however which was severely affected by strong wind attacks, the actual status of the genetic system must be judged to be in critical conditions
Prunus - The results obtained show significant differences in the level of genetic diversity, genetic differentiation and spatial genetic structure. In all cases the genetic differences among stages of different stands were significantly larger (20-40%) than the differences among stages of the same stand (< 15%).
A proportion of clones between 6% and 81% of all individuals was found to be present. Estimates of pollen and seed dispersal as well as appraisals of the proportion and spatial extension of vegetative regeneration for each stand were calculated. The mean pollen dispersal varied in the range of 45-162 m and the mean seed dispersal within 27-73 m. Among the stands the start of flowering differed by 10 days. Within a stand the flowering of most trees was found to overlap. A broad variation of tree densities was observed ranging from 5.3 adult trees per ha to 230 adult trees per ha. The distances of seed dispersal were found to be independent from of the tree densities. Pollen dispersal however was partly controlled by densities and even more by the extension of clonal groups. Due to the self-incompatibility system auto-fertilization was impossible for trees presenting the same clone.
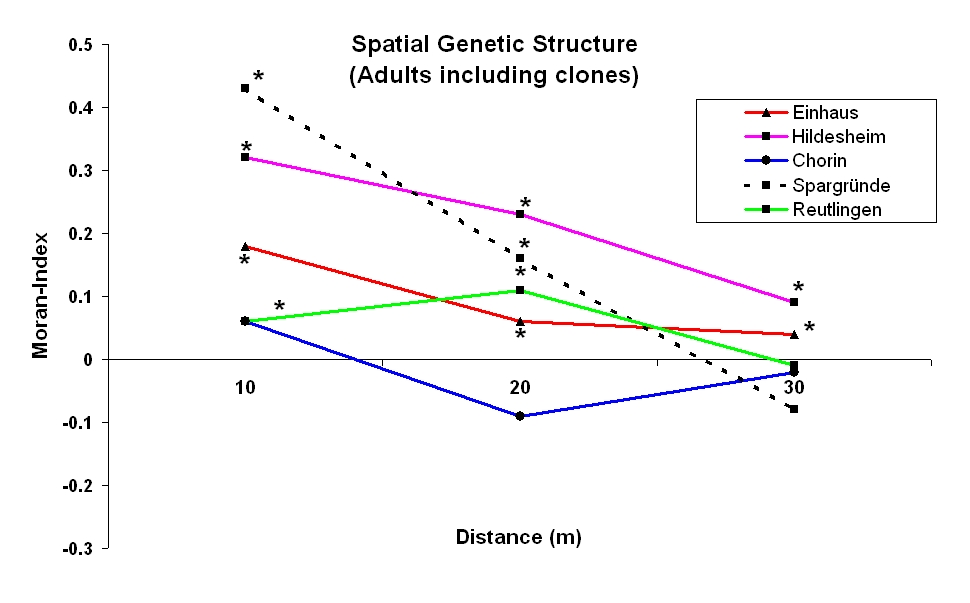
For the Moran’s index which gives information of the so-called spatial autocorrelation, significantly positive values in four out of the five plots under study were obtained for the first three distance classes up to 30 m (see Fig. 3). The bulk of these spatial genetic structures most probably results from the clones present.
Fig. 3 - Moran-index of the adult trees for four microsatellites as determined in the five wild cherry monitoring plots by including all trees growing within the plots; (*): significant on 95 % level.
The genotypes of the adult populations and the estimates of population-genetic processes were integrated into the simulation model Eco-Gene. The simulations predicted the dynamics of genetic structures and supported defining thresholds for critical changes in the genetic composition of the wild cherry populations. A set of 13 indicators and verifiers was tested according to the Concept of Genetic Monitoring to estimate the intactness of the genetic system of the five stands under study. For the indicator “level of genetic diversity” the effective numbers of genetic variants (alleles) in the three types of studied gene markers were used as verifiers. To verify the indicator “mating system”: (a) in accordance with Hardy-Weinberg proportions; (b) effective distances of pollen dispersal; (c) proportion of clones; (d) presence of family structures; (e) number of different S-alleles for the gene locus which controls the self-incompatibility of pollen; and (f) the germination percentage of seeds was taken into account. The indicator “genetic drift and directional change of genetic structure” was evaluated using the verifiers: (a) genetic differences among different ontogenetic stages of the same stand; (b) risk of hybridization with cultural cherry trees; and (c) population size. Like for beech each verifier got one of the three values -1, or Zero (0) or +1. The average of all verifiers was used to assess the overall situation of each population. This allowed a clear ranking among the stands concerning the quality, i.e., intactness of the genetic system.
Conclusions
In a holistic approach genetic monitoring includes genetic and phenological as well as physiological aspects of equal significance for characterizing the genetic system of tree populations. It is in fact an early warning and controlling system, because changes in the stand structure, vitality and natural regeneration often proceed with concomitant changes of the genetic system, which are usually not immediately observable. The results of the genetic monitoring supply information on the current state of the genetic system of the forest tree species under surveillance. Moreover, they are judged to be useful for estimating genetic implications of silvicultural measures; for developing practical strategies concerning the conservation of forest genetic resources; for integrating genetic aspects in the strategic utilization of different monitoring programmes as well as for optimizing the genetic management in view of the sustainable utilization of forest ecosystems.
The two pilot studies carried out gave first important practical experiences. Moreover, they reveal the necessity and urgency to initiate genetic monitoring promptly by considering different tree species. And finally genetic monitoring should be performed coordinated both on the national and the international level.
It cannot be answered presently whether or not all indicators examined deserve the same weighting. Further research in this context is absolutely necessary. With these case studies, the first proper monitoring of plots with a rich genetic data pool was initiated. The genetic monitoring of the plots presented here will be continued in the future.
Acknowledgement
The authors wish to thank all the members of the BLAG-Expertengruppe “Genetisches Monitoring”, especially Ute Tröber, Eva Cremer, Aikaterini Dounavi, Celine Jolivet, and Winfried Steiner as well as all other colleagues who contributed to the overall project with their target-oriented work and cooperative dedication. They also appreciate the competence and skilfulness of the lab personnel. And last but not least they are obliged to the Bundesamt für Landwirtschaft und Ernährung (BLE; Federal Agency for Agriculture and Food) in Bonn (Germany) for funding the projects (Az. 514-73.01/ 05BE003/1 and 05/BE003/2).
References
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Bayerisches Amt für forstliche Saat- und Pflanzenzucht, Forstamtsplatz1, D-83317 Teisendorf (Germany)
Forschungsanstalt für Waldökologie und Forstwirtschaft Rheinland-Pfalz, Schloss, D-67705 Trippstadt (Germany)
Institut für Forstgenetik des von Thünen-Instituts, Sieker Landstrasse 2, D-22927 Großhansdorf (Germany)
Landesforstanstalt Eberswalde, Fachbereich Waldentwicklung und Monitoring, Alfred-Möller-Strasse 1, D-16225 Eberswalde (Germany).
Corresponding author
Paper Info
Citation
Konnert M, Maurer W, Degen B, Kätzel R (2011). Genetic monitoring in forests - early warning and controlling system for ecosystemic changes. iForest 4: 77-81. - doi: 10.3832/ifor0571-004
Paper history
Received: May 18, 2010
Accepted: Sep 19, 2010
First online: Apr 05, 2011
Publication Date: Apr 05, 2011
Publication Time: 6.60 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2011
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 60597
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 49328
Abstract Page Views: 4280
PDF Downloads: 5162
Citation/Reference Downloads: 37
XML Downloads: 1790
Web Metrics
Days since publication: 5430
Overall contacts: 60597
Avg. contacts per week: 78.12
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Mar 2025)
Total number of cites (since 2011): 15
Average cites per year: 1.00
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Review Papers
Monitoring the effects of air pollution on forest condition in Europe: is crown defoliation an adequate indicator?
vol. 3, pp. 86-88 (online: 15 July 2010)
Research Articles
Effects of abiotic stress on gene transcription in European beech: ozone affects ethylene biosynthesis in saplings of Fagus sylvatica L.
vol. 2, pp. 114-118 (online: 10 June 2009)
Research Articles
Patterns of genetic diversity in European beech (Fagus sylvatica L.) at the eastern margins of its distribution range
vol. 10, pp. 916-922 (online: 10 December 2017)
Research Articles
Long-term monitoring of air pollution effects on selected forest ecosystems in the Bucegi-Piatra Craiului and Retezat Mountains, southern Carpathians (Romania)
vol. 4, pp. 49-60 (online: 05 April 2011)
Research Articles
Identification and molecular characterization of LTR and LINE retrotransposable elements in Fagus sylvatica L.
vol. 2, pp. 119-126 (online: 10 June 2009)
Research Articles
Variability of tolerance of Wild cherry clones to PEG-induced osmotic stress in vitro
vol. 15, pp. 265-272 (online: 25 July 2022)
Editorials
COST Action FP0903: “Research, monitoring and modelling in the study of climate change and air pollution impacts on forest ecosystems”
vol. 4, pp. 160-161 (online: 11 August 2011)
Research Articles
A fast screening approach for genetic tolerance to air pollution in Scots pine field tests
vol. 6, pp. 262-267 (online: 01 July 2013)
Commentaries & Perspectives
Clean air policy under the UNECE Convention on long-range transboundary air pollution: how are monitoring results “translated” to policy action
vol. 2, pp. 49-50 (online: 21 January 2009)
Research Articles
Tracing the acclimation of European beech (Fagus sylvatica L.) populations to climatic stress by analyzing the antioxidant system
vol. 14, pp. 95-103 (online: 01 March 2021)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







