
Tree encroachment dynamics in heathlands of north-west Italy: the fire regime hypothesis
iForest - Biogeosciences and Forestry, Volume 3, Issue 5, Pages 137-143 (2010)
doi: https://doi.org/10.3832/ifor0548-003
Published: Sep 27, 2010 - Copyright © 2010 SISEF
Research Articles
Abstract
Tree encroachment is one of the primary conservation issues in Calluna-heathlands, a priority habitat in Europe. Improving understanding of the ecological factors that trigger transitions to woodlands is key to developing strategies for heathlands management. The irrational use of fire has been recognized as one of the key factors that drives the loss of heathlands of north-west Italy. The effect of high frequency pastoral burning on the replacement of heathlands by grasslands has been documented by several studies. The relationship between fire and tree encroachment is less clear. The paper examines the effect of the fire regime on the encroachment of Populus tremula L. and Betula pendula Roth. in the heathland. The study was carried out at the Managed Nature Reserve of Vauda (7° 41’ E, 45° 13’ N), which includes one of the most valuable heathlands of north-western Italy. The experimental design consisted of analysing the age structure, dendrometric variables and the species composition of three aspen and birch stands, circular-shaped and isolated within the heathland matrix. From 1986 to 2009 all stands experienced the same fire regime due to pastoral burning. Wildfires of similar behaviour occurred in 1998, 2003 and 2008 and determined the stand structure observed in 2009. The results evidenced that fire acts as a catalyst not only for seedlings establishment, as previously documented, but also for woodland expansion in the heathland. After initial establishment, stands showed a concentric encroachment dynamic, mainly due to aspen root suckering after post-fire stem mortality, whose steps of expansion coincided with the return interval of wildfires. Moreover, aspen determined the loss of heathland characteristic species, whose relative abundances were inversely correlated to aspen density along a gradient from the stands centre to the surrounding heathland. The regulation of current burning practices by prescribed burning, integrated with rational grazing, presents the next research questions to be addressed.
Keywords
Tree encroachment, Heathland, Populus tremula, Conservation management, Prescribed burning
Introduction
Tree encroachment is one of the primary conservation issues in Calluna vulgaris (L.) Hull. dominated heathlands ([19]), a priority habitat (Habitats directive 92/43/EEC) and one of the most important cultural landscapes in Europe ([14]). Heathlands developed as a consequence of anthropogenic forest clearance and Calluna competition strategies ([23], [35], [26], [32], [6]). Furthermore, heathlands owe their continuance to forestry, pastoral or agricultural activities ([22]). Tree felling, rotational burning, grazing and harvesting the vegetation for fodder have been common characteristics of heathland management throughout their range of distribution ([41]).
Woodland encroachment that follows the loss of traditional management practices has been documented extensively for Atlantic heathlands of western and central Europe ([21], [28], [36], [27], [20]). In southern Europe, few studies have addressed this issue ([5], [7]), despite a range of concerns regarding tree encroachment. First, Calluna vulgaris reaches its ecological and geographical limits and its competitiveness is limited when faced with encroachment ([17], [18]). Second, the encroachment process occurs faster than in Atlantic regions ([19], [5], [7]), and, lastly, Calluna-heathlands are fragmented in vulnerable and restricted areas of high ecological value ([5], [31], [1], [7]).
In north-west Italy, Calluna-heathlands are secondary coenoses that colonized acidic and oligotrophic soils where Querco-Fagetea forests have been harvested or burned, and were used as pastures or to harvest fodder ([39], [1], [7]). Until the post-WWI period, heathlands covered extensive areas ([30]), but today they are limited to isolated patches within the agricultural and urban mosaic. The majority of remaining heathlands have been designated as “Managed Nature Reserves” (MNR) in order to safeguard them from deterioration and preserve their values through management ([39], [37]). Despite this protection regime, several studies have observed the loss of the heathland habitat and an increase of woodlands throughout north-west Italy ([33], [16], [1], [7]). Improving understanding of the ecological factors that trigger transitions to woodlands is the key to developing strategies for heathlands conservation management.
The irrational use of pastoral burning has been recognized as one of the key factors that drives the loss of Calluna-heathlands at the MNR of Vauda, which include one of the most valuable heathlands of north-west Italy ([29], [3], [7], [25]). Uncontrolled fires every 1-5 years characterize the fire regime of extensive heathland areas within the Reserve ([29], [7]). Local shepherds ignite the fire in the driest months of late winter, using the föhn wind ([10]) to spread the fire over large areas from the north-west to the south-east side of the Reserve, in order to renew pastures and graze their cattle in spring.
The effect of high frequency burning on the replacement of Calluna-heathlands by grasslands has been documented by several studies and authors agree on the ecological processes involved ([29], [3], [7], [25]).
The relationship between the current fire regime and tree encroachment is less clear. A study involving repeated aerial photographs of the MNR of Vauda (1964-2004 period) showed evidence of the gradual loss of the heathland habitat mainly due to an increase in Populus tremula L. (aspen) and Betula pendula Roth (birch) woodlands ([33]). Moreover, the rate of tree encroachment accelerated in the last two decades, with an increase of 476% in cover of young aspen and birch stands against the loss of 44% of the heathland ([33]).
Is this dramatic expansion of woodland related to the current fire regime?
According to Mugion ([29]), the elimination of the Calluna vulgaris by frequent burning depletes the allelopathic substances produced by its roots, thus opening the way for tree encroachment. Borghesio ([7]) reports that frequent, low intensity fires appear to enhance woodland expansion: fire stimulates seed germination and seedling establishment of Betula pendula by providing patches of bare soil where competition is low. Moreover, in frequently burned areas, fire intensity is low as a consequence of periodic fuel reduction; consequently the rapid growth of birch seedlings enhances their chances of reaching a stage where they are safe from low intensity fires.
In view of the importance of the heathland habitat in north-west Italy, a better understanding of the relationship between woodland encroachment and the current pastoral burning practices is the key to define suitable management programs for heathland conservation. This paper examines the effect of the fire regime on the age structure and expansion patterns of Populus tremula and Betula pendula stands in the Calluna-heathland habitat. Finally, it discusses the management implications.
Materials and methods
The MNR of Vauda is located 20 km N-NE of Torino (7° 41’ E, 45° 13’ N) and covers an area of 2635 ha at an altitude ranging from 240 to 480 m. The Reserve is located on a fluvio-glacial terrace dated to the Mindel glaciation (c. 400.000 years ago). Soils are ancient and leached, with fairly low pH (4.8 ± 0.1 - [7]). The climate is transitional between continental and suboceanic, characterized by a yearly average precipitation of about 1.130 mm, 305 mm of which accumulates during summer. The mean annual temperature is 11.8°C, with monthly means ranging from 1.4°C in January to 22.1°C in July. The driest month is March with, on average, 35 mm of rain and 0.3 days of snow ([25]).
The study site was located in the central sector of the MNR of Vauda (Fig. 1a), an area with one of the largest heathlands in the Reserve ([29]). Additionally, over the few last decades the area has been affected by frequent wildfires due to pastoral burning ([7]), and has experienced major increases in woodlands ([33]).
Fig. 1 - a) Map of the Managed Nature Reserve of Vauda showing the Reserve boundaries (green line), the location of the study site (black line) and the areas burnt within 1996 and 2009 (red area). (b) Quickbird satellite image of the study site dating back to 2004 ([33]; IPLA S.p.A. ©) showing several isolated forest stands, circular-shaped within the heathland matrix.
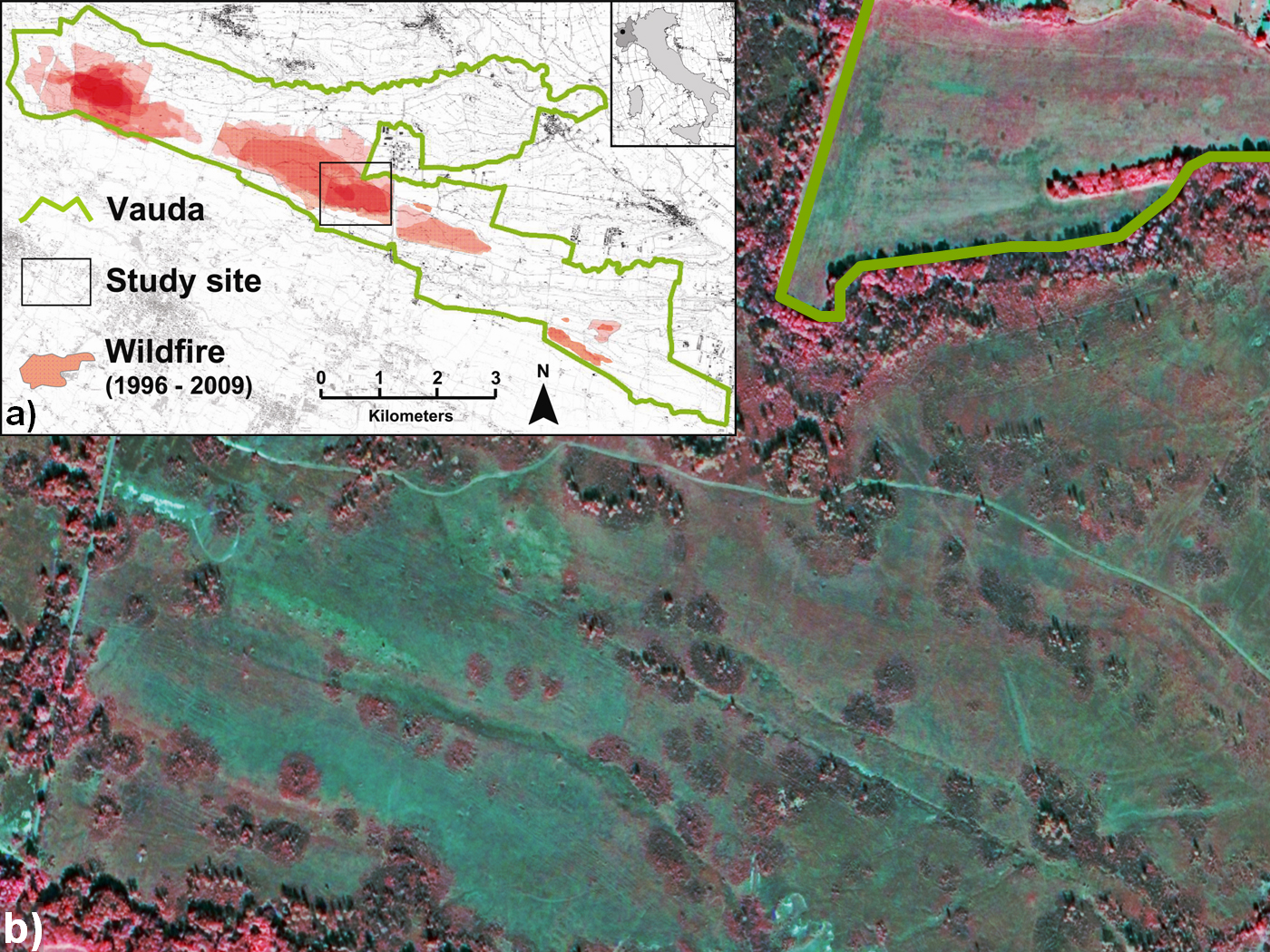
Analysis of a Quickbird satellite image of the study area from 2004 (Fig. 1b) showed many isolated, circular shaped stands within the heathland matrix. These stands, mainly constituted by Populus tremula and Betula pendula, were not present at the end of 1970s ([33]). Consequently, they developed during the period of major expansion of woodlands, and their structure could represent a key to better understanding the encroachment dynamics within the heathland in relation to the fire regime of the area.
The experimental design consisted of analysing the age structure, dendrometric variables and the species composition of three stands which have been selected on the basis of the following characteristics: i) an area of at least 0.2 ha, so as to be considered a forest stand according to the Italian and Regional normative (D.lgs. 227/2001 and L.R. 4/2009 respectively); ii) the stands had to have a similar size and be surrounded by heathlands on all sides; and iii) all stands had to be affected by the same fire regime.
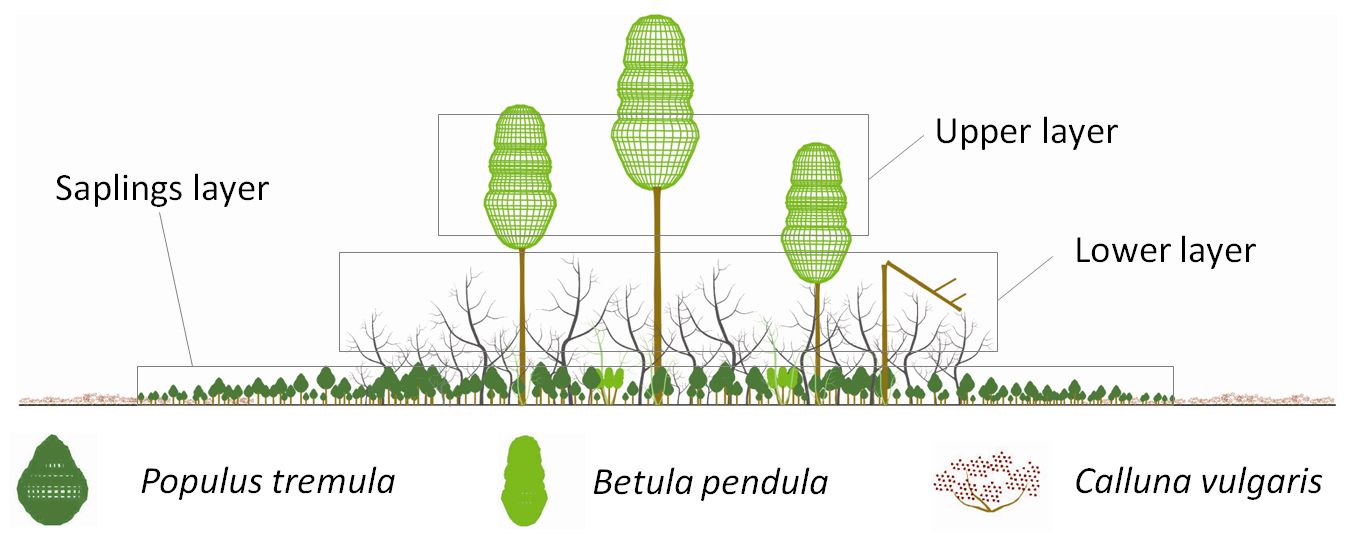
The age structure and dendrometric variables of each stand were characterized in spring 2009. All stands had a multi-layered structure characterized by three distinct layers (Fig. 2): a low-density upper-layer mainly comprised of Betula pendula distributed as individuals with evident fire scars on the trunk; a lower-layer made up of a thick stand of Populus tremula and Betula pendula, which were all dead; finally, a high density layer of saplings, mainly of Populus tremula.
Fig. 2 - Illustration showing the multi-layered vertical structure of the studied stands as observed in spring 2009: the upper-layer was comprised only of birch distributed as individuals; the lower-layer was made up of a thick stand of aspen and birch, which were all dead; the saplings-layer was constituted mainly by a high density layer of aspen.
The sampling method was different for each layer:
- Upper-layer: species, diameter at 1.3 m, tree height, age (by counting rings in cores extracted from the scar area and the non- scarred area of the trunk), height of fire scars, crown mortality (according to 4 classes of crown damage expressed as percentage of the crown volume: 0-33%; 33-66%; 66-99%; 100% or dead) and the georeferenced position (recorded with a GPS) were collected for each individual of the stand;
- Lower-layer: species, diameter at 1.3 m, height and crown mortality were measured for all of the individuals within 12.56 m2 circular sampling plots randomly distributed within the stand (5 per stand). Age was measured on a sub-sample of 10 individuals per sampling unit by counting rings of a section cut at the stem base. The perimeter of the area on which the lower-layer persisted was georeferenced with a GPS.
- Saplings-layer: species, diameter at the root-collar, height and crown mortality were measured for all of the individuals within a 3.14 m2 circular sampling unit randomly located within the stand (10 per stand). Age was measured on a sub-sample of 10 individuals per sampling unit. The perimeter of the sapling layer was georeferenced with a GPS.
Variable means were compared with the analysis of variance including the year in which wildfires occurred and the stand as fixed factors. Significant differences were tested with the Bonferroni post-hoc range test (P < 0.05). Data were examined for homogeneity of variance (Levene test).
The grass and shrub species of the understory (including tree saplings) were surveyed using the vertical point quadrat method ([12]) along 30-m transects (4 per stand). The transects had origins in the centre of the stand and faced the four cardinal points, in order to assess species abundance along a gradient from the interior of the stand to the surrounding heathland on all sides. In each transect, at each 20-cm interval, species touching a steel needle were identified and recorded. Records were divided into sets corresponding to segments of 5 m in length, which divided each transect into 6 portions progressively distanced from the stand centre. For each segment the absolute number of point intercepts of every species (SFi) was calculated. Species relative abundance (SCi) for each segment was then calculated according to the following equation ([12]- eqn. 1):
The SCi of each species was finally studied in relation to the distance of the segment from the origin (centre of the stand) by linear regression analyses. Correlation analysis was used to test competition between species.
All analyses were performed using the SPSS v.16.0 statistical package ([40]).
Results and discussion
The fire regime of the study area was determined for the last two decades. According to the wildfire statistics of the Corpo Forestale dello Stato (1986-2009 period) and wildfire perimeters documented by Borghesio ([7]) - integrated with the geo-referenced fire perimeters provided by the Land Managers of the MNR of Vauda for the period 1996-2009 -, from 1986 to 2009 the selected stands were impacted by three wildfires. After a relatively long period with an absence of fire, the first wildfire dated back to 08/03/1998 and covered an area of 270 ha; the second one occurred on 27/03/2003 with an area of 122 ha; and the third one burned on 24/03/2008 with an area of 222 ha.
The fire scar analysis confirmed that all stands experienced the same fire regime. In 2009, the majority of birch individuals of the upper layer showed two scars overlapping on the south-east side of the trunk (Fig. 3). The difference in the number of tree rings between cores extracted from the scar area and the non-scarred area ranged from 10 to 11 years. Consequently, the deeper scar corresponds to the 1998 fire, as reported by wildfire statistics of the study area. The overlapped scar is probably due to the 2003 wildfire, but it was not possible to determine by tree ring counting. The charred bark on the south-east side of the trunk verified the 2008 wildfire. Average (± SE) scar heights of the 1998 and 2003 wildfires were 1.35 ± 0.05 m and 1.44 ± 0.07, respectively, and were not significantly different among stands (F[2.41] = 0.137, P < 0.872) and years (F[1.41] = 2.466, P < 0.124). The average height of the charred bark after the 2008 wildfire was 1.60 ± 0.04. Consequently, the three wildfires behaved similarly: they were all winter fires occurring in the driest month of March in heath fuels; they all spread with the föhn wind from north-west to south-east; and finally they manifested a similar severity on the trunk as evidenced by fire-scar heights.
Fig. 3 - Fire scars on a birch trunk of the upper-layer which are evidence of the fire regime of the area. The image shows two overlapped scars which correspond respectively to the 1998 and 2003 wildfires, while the charred bark is due to the 2008 wildfire.

All stands showed the same age structure and dendrometric characteristics. Average values of variables observed in spring 2009, together with the results of the analysis of variance, are reported for each layer in Tab. 1. No significant differences were found among stands for most of the measured variables. Consequently, we can state that all stands belong to the same population and are representative of the encroachment dynamic in relation to the fire regime of the area.
Tab. 1 - Average (±SE) value measured in spring 2009 for the following variables: tree density, species composition (Bp: Betula pendula, Pt: Populus tremula), crown damage (Class 1: live, Class 4: dead), age, diameter, tree height and area of the upper, lower and saplings layers. Results of the analysis of variance which tested significant differences among stands are reported below the variable mean. (*): P<0.05; n.s.: no significant differences).
| Layer | Upper | Lower | Saplings |
|---|---|---|---|
| Density (stem ha-1) | 37 ± 8 - |
51.106 ± 6.415 (F2.12 = 2.29; P < 0.18 n.s.) |
145.185 ± 12.157 (F2.27 = 0.46; P < 0.64 n.s.) |
| Species composition (%) | 100% Bp 0% Pt |
13% Bp 87% Pt |
6% Bp 94% Pt |
| Crown mortality (4 classes) |
Class 1: 20%; Class 2: 20% Class 3: 24%; Class 4: 36% |
Class 1: 0%; Class 2: 0% Class 3: 0%; Class 4: 100% |
Class 1: 100%; Class 2: 0% Class 3: 0%; Class 4: 0% |
| Age (yr) | 21 ± 2.4 (F2.27 = 1.76; P < 0.19 n.s.) |
4.7 ± 0.1 (F2.12 = 0.25; P < 0.78 n.s.) |
1 - |
| Diameter (cm) | 14.9 ± 0.7 (F2.27 = 0.27; P < 0.76 n.s.) |
2.1 ± 0.1 (F2.12 = 0.50; P < 0.62 n.s.) |
0.9 ± 0.06 (F2.27 = 0.58; P < 0.57 n.s.) |
| Height (m) | 9.5 ± 0.9 (F2.27 = 3.85; P < 0.34*) |
2.3 ± 0.1 (F2.12 = 0.90; P < 0.43 n.s.) |
0.61 ± 0.04 (F2.27 = 0.12; P < 0.89 n.s.) |
| Area (m2) | - | 1721 ± 188 | 2645 ± 246 |
The upper layer was made up of only birch individuals with an average age of 21 years; consequently birches established at the end of 1980s. It is difficult to determine if their origin was gamic or agamic but the distribution of individuals could indicate a seed origin. Several authors report that birch establishes on bare soils, as the ability of seedlings to penetrate the canopy of Calluna is low, whereas in open conditions birch is at an advantage and can compete effectively with other species ([21], [4], [34], [27]). Recently burned areas are ideal for germination of birch seeds and the establishment of seedlings: ash promotes seed germination and increases root growth during the most sensitive period of seedlings development, thus promoting their survival ([18], [4]). Moreover, burning may remove allelophatic effects of the Calluna ([29], [6]). Consequently, we could suppose that birch individuals of the upper-layer established within the heathland as a consequence of a disturbance in the mid-1980s. This disturbance event could be a wildfire, but no fire statistics are available for the study site before 1986.
What is certain is that most birch individuals of the upper-layer resisted the 1998, 2003 and 2008 wildfires. According to the crown mortality observed in 2009 (Tab. 1), only 36% of birch trees were dead (Class 4), whereas 20% of the individuals did not show crown mortality (Class 1). Betula pendula, in fact, has a moderate resistance to fire ([34]). Preliminary results of a field fire experiment carried out at the MNR of Vauda ([2]), which studied the effects of fire behaviour on the probability of tree mortality vs. stem diameter, showed that birch stem mortality decreases significantly for individuals with a diameter larger than 6 cm, both after fire fronts of low intensity (< 500 kW m-1) and of moderate intensity (500-2000 kW m-1). Consequently, we could suppose that birch individuals of the upper layer, in the absence of fire from their establishment until 1998, managed to grow large enough that they were able to survive consecutive wildfires of moderate intensity in 1998, 2003 and 2008.
The lower-layer was made up of 87% Populus tremula and 13% Betula pendula with an average age of 4.7 years. All individuals were dead (Tab. 1). It is not clear if aspen established at the same time as birch, or entered the stand after birch was already present. It is known that fire promotes the germination of aspen seeds and early seedling development ([24]). Consequently, it could have established in the mid-1980s, after the same disturbance event that allowed birch establishment. We could also suppose that birches modified the environment, thus providing suitable sites for aspen ([27]). In any case, neither snags of aspen nor living individuals dated before 2003 were found in the stand. Consequently, aspen stems, which constituted the early stand, did not survive the 1998 or 2003 wildfires, and their snags were eliminated both by wood decay and by combustion. Nevertheless, the aspen capacity to regenerate asexually by root suckering in a disturbed environment has been well documented ([42], [38], [24], [11]). After a fire, when apical dominance is reduced as a consequence of post-fire stem mortality, sucker-stimulating cytokinins increase in the roots where buds are protected from heat by the soil stratum, resulting in high sprout capability ([8]). This sprouting gives origin to thick coetaneous stands ([38]). The lower-layer observed in 2009 was in fact an even-aged population with an average density equal to 51,106 stem ha-1. Additionally, very little variability was found in stem diameter and height (Tab. 1). As a consequence of the stem mortality induced by the 2003 wildfire to the pre-fire lower-layer, we could suppose that birch in the lower-layer generated from stump suckering, and that aspen generated from root suckering. Subsequently, suckers grew until 2008 when a new wildfire induced 100% stem mortality, as individuals did not yet have the size that could have conferred them resistance traits. This result confirms previous studies which document 100% stem mortality for birch and aspen diameters lower than 6 cm ([8], [2]).
As a consequence of the lower-layer stem mortality induced by the 2008 wildfire, a new generation of suckers sprouted. The saplings-layer that originated showed a substantial increase in density up to 145 000 stem ha-1, 94% of which were aspen shoots of one year (Tab. 1).
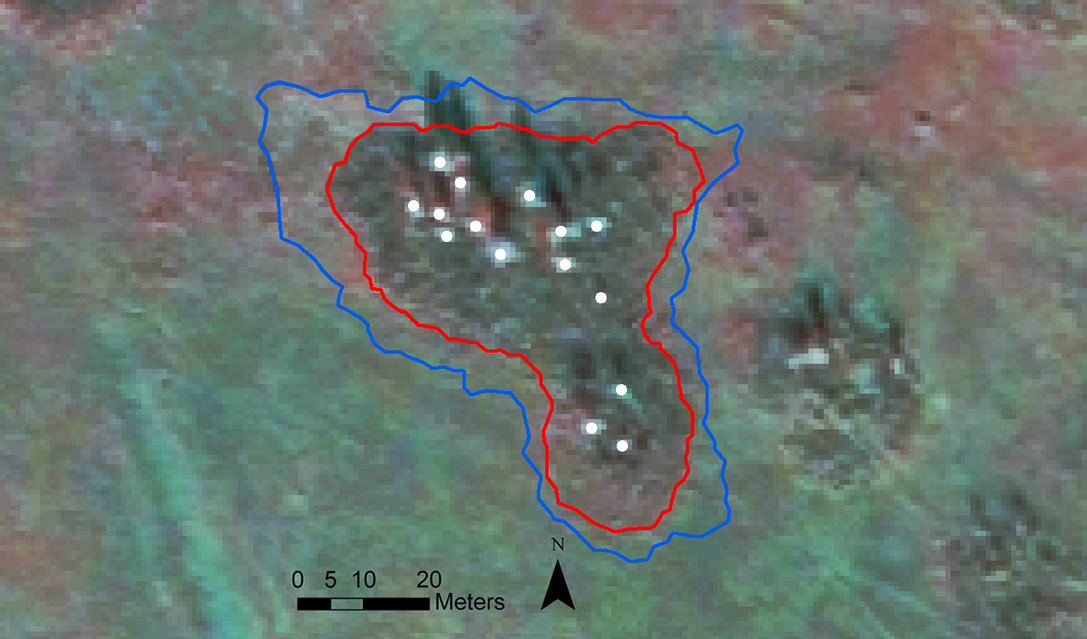
An interesting result was obtained by analysing the horizontal structure of stands by superimposing the perimeters of stand layers in a GIS environment. In all stands, all the individuals of the upper-layer were included within the area of the lower-layer, which in turn was included within the area of the saplings-layer (Fig. 4). The average increase in area from the lower-layer to the saplings- layer was 154 ± 26 %. Consequently, stands showed a concentric encroachment dynamic, mainly due to aspen root suckering, whose steps of expansion coincided with the return intervals of wildfires.
Fig. 4 - Quickbird satellite image of one of the studied stands in 2004, to which it has been superimposed the position of birch individuals of the upper-layer (white dots), which are all included within the perimeter of the lower-layer (red line) which in turn is included within the perimeter of the saplings-layer (blue line).
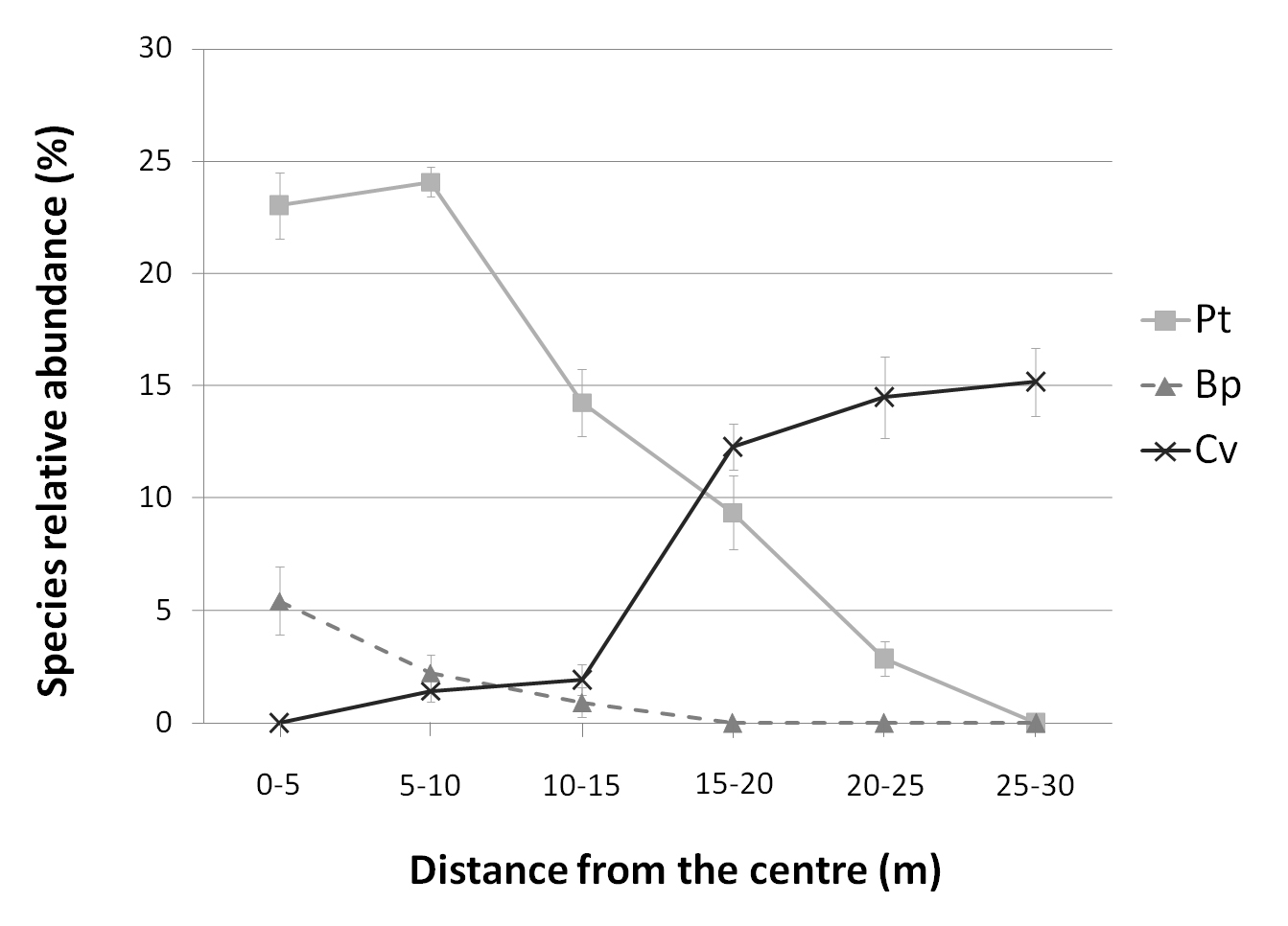
The species composition of the grass and shrub understory showed substantial changes from the interior of the stand to the surrounding heathland. Linear regression analysis showed a significant decrease in species relative abundance (SCi) of aspen (P < 0.01; R2 = 0.84) and birch (P < 0.01; R2 = 0.34), and a significant increase of Calluna (P < 0.01; R2 = 0.72), at increasing distances from the stand centre (Fig. 5). The Calluna distribution showed a marked rise at a distance between 15-20 m from the centre, which corresponds to the new encroached area by aspen suckers after the 2008 wildfire. At larger distances, it stabilizes around 12%, which is in accordance with the SCi observed at the second vegetative season after experimental fires conducted in the MNR of Vauda to test Calluna regeneration rates in absence of tree encroachment ([2]).
Fig. 5 - Species relative abundance (SCi) along a gradient from the interior of the stand to the surrounding heathland habitat in 2009. Graphs represents average SCi (±SE) for progressive distances from the stand centre of the following species: Pt: Populus tremula; Bp: Betula pendula; Cv: Calluna vulgaris.
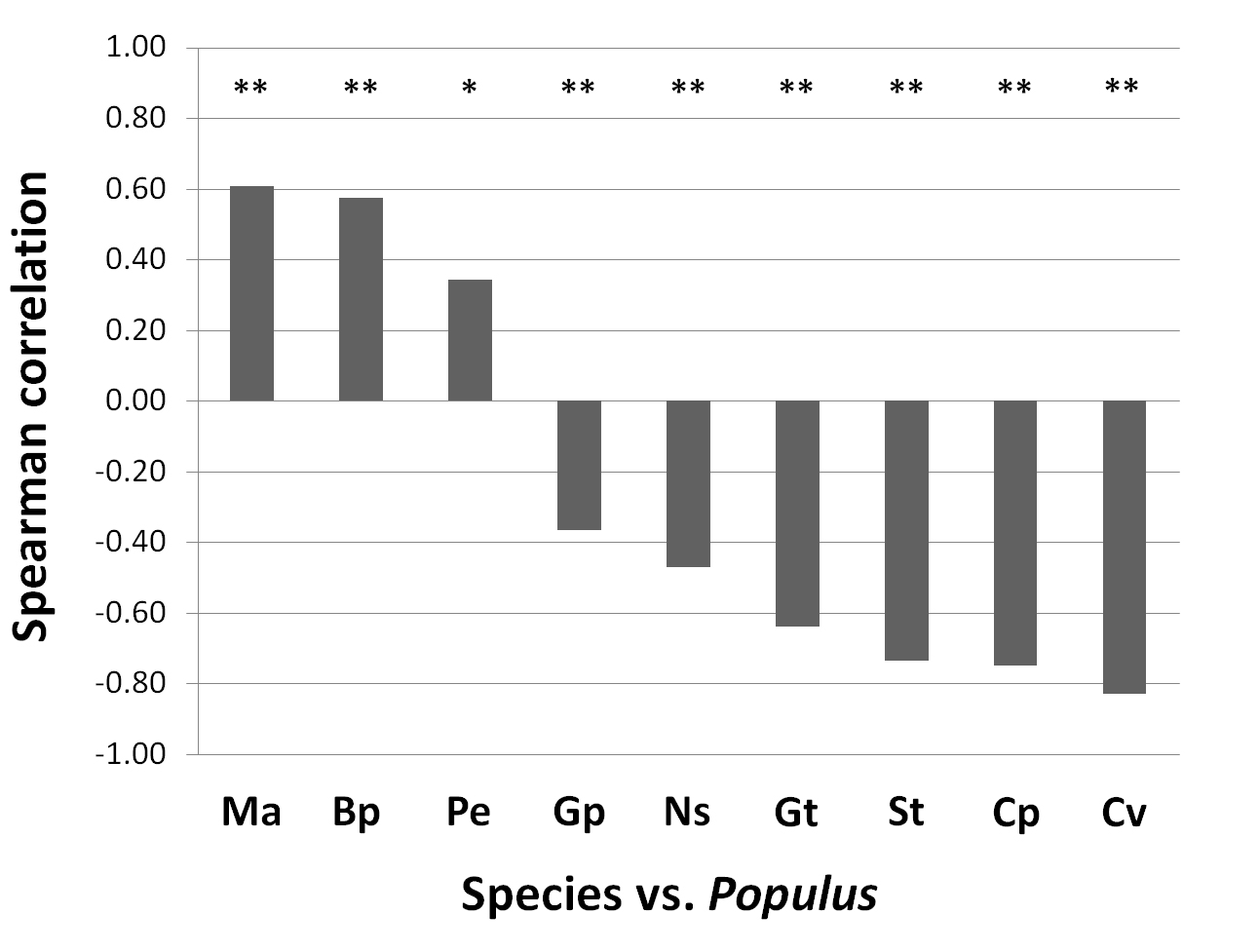
The marked difference in species relative abundance between the interior of the stand and the surrounding heathland was related to aspen density. The Spearman correlation coefficients between the SCi of heathland species vs. aspen are reported (Fig. 6). Molinia arundinacea was positively correlated to aspen, denoting its strong competitiveness under the tree cover and its marked resilience to fire ([9]). By contrast, Gentiana pneumonanthe L., Nardus stricta L., Genista tinctoria L., Serratula tinctoria L., Carex panicea L. and Calluna vulgaris, which characterise the heathland habitat of this portion of the MNR of Vauda ([29], [33]), were negatively correlated to aspen.
Fig. 6 - Spearman rank correlation coefficients and significance (**: P < 0.01; *: P < 0.1) between the SCi of species which characterize the heathland habitat and the SCi of aspen assessed along transects (N = 72 estimates). Species abbreviations means: Ma: Molinia arundinacea, Bp: Betula pendula, Pe: Potentilla erecta, Gp: Gentiana pneumonanthe, Ns: Nardus stricta, Gt: Genista tinctoria, St: Serratula tinctoria, Cp: Carex panacea, Cv: Calluna vulgaris.
Conclusions
The present study enabled a deeper understanding of the ecological processes involved in birch and aspen encroachment dynamics in Calluna-heathlands of north-west Italy. Fire is one of the main determinants that trigger transition towards woodlands. Fire provides suitable sites for birch and aspen seed germination and seedling establishment by removing the vegetation cover and depleting allelophatic compounds, as previously observed by Borghesio ([7]) and Mugion ([29]). After initial establishment, woodland encroachment is mainly due to aspen agamic regeneration by root suckering after post-fire stem mortality, as evidenced by the present study. Moreover, aspen is the main species that determines the loss of heathland characteristic species, as previously observed by several authors ([29], [33], [39], [7]).
Consequently, fire acts as a catalyst not only for tree establishment, but also for tree stand expansion in the heathland. After each fire event, isolated stands enlarge their area until they merge together. This expansion dynamic also takes place at the border between the heathland habitat and the surrounding birch and aspen forests, thus determining the enclosure of the remaining patches of the heathland. This thesis could explain the increasing rate of transition from heathlands to woodlands, which characterised recent landscape dynamics in the MNR of Vauda ([33], [7]) in correspondence with the increase in fire frequency due to the irrational use of fire by shepherds over last two decades.
Consequently, the current pastoral practices, characterized by the frequent use of fire with a return interval from 1 to 5 years over large areas, and the decrease in grazing stock rate ([7]), are primarily responsible for the loss of the heathland habitat. The reintroduction of traditional burning practices by prescribed burning ([15]), integrated with rational grazing has been applied in many heathland management programs throughout Europe ([22], [20], [13]), and testing its feasibility for tree control and heathland conservation management in north-west Italy is currently the object of experimental studies ([3], [25]). The debate over burning prescriptions, namely the fire-treatment frequency, season and behaviour presents the next research questions to be addressed.
Acknowledgments
The authors thanks Pilan Emanuele, for his contribution to the field work, and the land managers of the Managed Nature Reserve of Vauda, for providing information about the fire regime of the area.
References
Gscholar
CrossRef | Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Gscholar
Authors’ Info
Authors’ Affiliation
Corresponding author
Paper Info
Citation
Ascoli D, Bovio G (2010). Tree encroachment dynamics in heathlands of north-west Italy: the fire regime hypothesis. iForest 3: 137-143. - doi: 10.3832/ifor0548-003
Paper history
Received: May 03, 2010
Accepted: May 28, 2010
First online: Sep 27, 2010
Publication Date: Sep 27, 2010
Publication Time: 4.07 months
Copyright Information
© SISEF - The Italian Society of Silviculture and Forest Ecology 2010
Open Access
This article is distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Web Metrics
Breakdown by View Type
Article Usage
Total Article Views: 50312
(from publication date up to now)
Breakdown by View Type
HTML Page Views: 43084
Abstract Page Views: 2571
PDF Downloads: 3421
Citation/Reference Downloads: 30
XML Downloads: 1206
Web Metrics
Days since publication: 4960
Overall contacts: 50312
Avg. contacts per week: 71.00
Article Citations
Article citations are based on data periodically collected from the Clarivate Web of Science web site
(last update: Feb 2023)
Total number of cites (since 2010): 21
Average cites per year: 1.50
Publication Metrics
by Dimensions ©
Articles citing this article
List of the papers citing this article based on CrossRef Cited-by.
Related Contents
iForest Similar Articles
Research Articles
Clonal structure and dynamics of peripheral Populus tremula L. populations
vol. 7, pp. 140-149 (online: 13 January 2014)
Review Papers
Prescribed burning in Italy: issues, advances and challenges
vol. 6, pp. 79-89 (online: 07 February 2013)
Research Articles
Near zero mortality in juvenile Pinus hartwegii Lindl. after a prescribed burn and comparison with mortality after a wildfire
vol. 12, pp. 397-402 (online: 31 July 2019)
Technical Notes
Goat grazing as a wildfire prevention tool: a basic review
vol. 7, pp. 260-268 (online: 26 March 2014)
Research Articles
Short- and long-term natural regeneration after windthrow disturbances in Norway spruce forests in Bulgaria
vol. 11, pp. 675-684 (online: 23 October 2018)
Research Articles
Soil stoichiometry modulates effects of shrub encroachment on soil carbon concentration and stock in a subalpine grassland
vol. 13, pp. 65-72 (online: 07 February 2020)
Research Articles
A state-and-transition approach to alpine grasslands under abandonment
vol. 3, pp. 44-51 (online: 02 March 2010)
Research Articles
Variation of major elements and heavy metals occurrence in hybrid aspen (Populus tremuloides Michx. × P. tremula L.) tree rings in marginal land
vol. 13, pp. 24-32 (online: 15 January 2020)
Research Articles
Post-fire effects and short-term regeneration dynamics following high-severity crown fires in a Mediterranean forest
vol. 5, pp. 93-100 (online: 30 May 2012)
Research Articles
Changes in vegetation diversity and composition following livestock removal along an upland elevational gradient
vol. 8, pp. 582-589 (online: 22 April 2015)
iForest Database Search
Search By Author
Search By Keyword
Google Scholar Search
Citing Articles
Search By Author
Search By Keywords
PubMed Search
Search By Author
Search By Keyword







